
Бактерии
| Бактерии Период: архаик–денес 3500–0 милиони години | |
|---|---|
.jpg?lang=mk)
| |
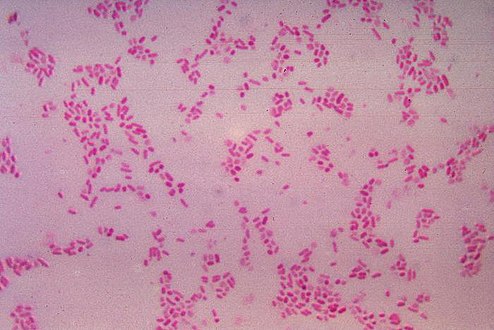
| Escherichia coli | |
| Научна класификација [ у ] | |
| Домен: | Бактерии Woese et al. 1990 |
| Синоними | |
| |
Бактерии (науч. Bacteria) — голем домен на прокариотски микроорганизми. Типично се неколку микрометри во должина и се среќаваат во голем број на облици, вклучувајќи сфери, стапчиња и спирали. Бактериите се едни од најстарите животни форми на планетата и може да се најдат во скоро сите хабитати. Тие ги населуваат океаните, копното, врелите извори, екстремно кисели води и радиоактивни отпади,[2] а може да се најдат дури и на големите длабочини на Земјината кора. Тие исто така живеат во сложените повеќеклеточни организми како што се животните и растенијата, а со нив формираат симбиотски или паразитски односи. Повеќето бактерии сè уште не се одликувани, а само околу половина од сите бактериски колена имаат видови кои може да се огледуваат во лабораторија.[3] Науката за бактерии се нарекува бактериологија и претставува гранка на микробиологијата.[4]
Во еден грам почва има околу 40 милиони бактериски клетки, а во милилитар слатка вода има околу милион бактериски клетки. На планетата Земја има околу 5×1030 бактерии.[5] Ова формира бактериска биомаса која е поголема од растителната и животинската биомаса заедно.[6] Бактериите се неопходни за многу фази од циклусот на елементите во биосферата, како што се врзувањето на јаглеродот и азотот од атмосферата. Циклусот на елементите исто така вклучува разградување на телата на мртвите организми, а бактериите се најодговорни за фазата на гниење на овој процес.[7] Во биолошките заедници кои живеат околу хидротермалните извори или студените извори, екстремофилните бактерии ги обезбедуваат виталните хранливи материи кои се неопходни да се одржи животот во овие средини; како што е искористувањето на метан и сулфурводород за продукција на енергија. Во 2013 година била објавена студија според која бактериите може да живеат во Маријанскиот Ров, кој се наоѓа на длабочина од 11 километри, што е најдлабокиот познат регион на океаните.[8][9] Други истражувачи објавиле резултати според кои бактериите може да живеат внатре во карпи кои се наоѓаат 580 метри под морското дно.[10] Верувањето дека бактериските клетки во човековото тело се побројни од човечките клетки во однос 10:1 е побиено. Има околу 39 трилиони бактериски клетки во човековата микрофлора, при што како стандард се зема маж со 170 сантиметри висина и 70 килограми телесна тежина, додека вкупниот број на човековите клетки е околу 30 трилиони. Ова значи дека иако бактериските клетки се побројни, тие се побројни за 30%, а не за 900% во однос на човековите клетки.[11] Најголемиот број на бактериски клетки кај човекот се наоѓаат во цревната флора, а втори по бројност се бактериите на кожата.[12] Најголемиот дел на бактериите кои живеат во човековото тело се сосема безопасни, а тие што живеат во цревната флора се дури и многу корисни. Меѓутоа неколку видови на бактерии се патогени и предизвикуваат инфективни заболувања како што се: чума, лепра, антракс, сифилис, колера, туберкулоза итн. Најсмртоносните бактериски заболувања се дишните индекции, од кои на прво место е туберкулозата која убива околу 2 милиона луѓе годишно главно во потсахарска Африка.[13] Во развиените земји за третман на бактериските инфекции често се користат антибиотици, кои исто така се употребуваат и во сточарството поради што резистенцијата кон антибиотици е во пораст. Во индустријата бактериите се користат за третман на отпадни води, за разградување на истурена нафта; во прехранбената индустрија за производство на јогурт, сирење и други производи на ферментација; како и во биотехнологијата за производство на антибиотици и разни други хемикалии.[14][15]
Порано бактериите се сметале за растенија и се класифицирале во класата Schizomycetes. Денес, бактериите се класифицираат како прокариотски организми кои, за разлика од животинските, растителните и другите еукариотски организми, не поседуваат клеточно јадро и немаат мембрански органели. Иако традиционално терминот „бактерии“ се користел за сите прокариоти, научната класификација е променета со почетокот на деведесеттите години од 20 век, по откритието дека археите се посебна група на прокариоти, кои заедно со бактериите еволуирале од заеднички предок. Денес бактериите и археите се класифицираат во два одделни домени на животот, а третиот го чинат еукариотите.[16]
Потекло на поимот
[уреди | уреди извор]Зборот бактерија потекнува од новолатинскиот bacterium, кој е латинизирана форма од грчкиот збор βακτήριον (bakterion),[17] што претставува деминутив на βακτηρία (bakteria), што во превод значи „стап“,[18] бидејќи првите видови кои биле откриени имале стапчеста форма.[19]
Потекло и рана еволуција
[уреди | уреди извор]Предокот на денешните бактерии бил едноклеточен микроорганизам и бил еден од првите живи форми кои се појавиле на Земјата пред околу 4 билиони години. Во следните 3 билиони години доминантните живи организми на Земјата биле едноставни прокариоти, како што се денес бактериите и археите.[20][21] Иако постојат бактериски фосили од архајскиот период, како што се строматолитите, недостатокот на одредена морфологија оневозможува тие да се искористат за проучување на историјата на бактериската еволуција, или да се прецизира времето на потекло на одреден бактериски вид. Меѓутоа во поново време, секвенционирањето на геномите на илјадници видови на организми од сите три домени на животот, овозможило да резултатите од овие истражувања се користат во филогенетиката.[22][23] Најблискиот древен заеднички предок на бактериите и археите веројатно бил хипертермофилен организам и живеел пред околу 2,5-3,2 билиони години.[24][25] Бактериите, исто така, влијаеле врз еволуцијата на еукариотите и археите. Денешните еукариоти се резултат на ендосимбиотска асоцијација помеѓу предокот на еукариотите (т.н. протоеукариот) и алфапротеобактерија, која после навлегување во цитоплазмата на протоеукариотот еволуирала во денешните еукариотски органели, како што се митохондриите, а според некои теории и хидрогенозомите.[26][27] Подоцна некои еукариоти, коишто веќе содржеле митохондрии, стапиле во секундарна ендосимбиотска асоцијација со цијанобактерија, што довело до еволуција на хлоропластите во денешните алги и растенија.[28]
Морфологија
[уреди | уреди извор]
Морфологијата на бактериите покажува голем диверзитет на форми и величини. Бактериските клетки се околу десетина од големината на еукариотските клетки и долги се околу 0,5 - 5,0 микрометри, но има и видови кои се видливи со голо око, како на пример Thiomargarita namibiensis, која е долга околу 0,5 милиметри,[29] и Epulopiscium fishelsoni која достигнува до 0,7 милиметри должина.[30] Најмалите бактерии се членови на родот Mycoplasma, кои имаат должина од само 0,3 микрометри, што претставува величина на најголемите вируси.[31] Некои бактерии можат да бидат уште помали, но овие т.н. ултрамикробактерии сѐ уште не се доволно проучени.[32]
Најголем број бактериски видови се или сферични по форма, т.н. коки (еднина: coccus, од грчкиот збор kókkos, што значи зрно или семе) или се со стапчеста форма, т.н. бацили (еднина: bacillus, од латинскиот збор baculus, што значи стап). Некои бактерии наречени вибрио имаат форма на искривено стапче или запирка; други пак имаат спирална форма и се наречени спирила, а пак други може да се значително свиени и се нарекуваат спирохети. Опишани се и помал број на други невообичаени форми, како на пример ѕвездовидни бактерии.[33] Бактерискиот клеточен ѕид и цитоскелет го одредуваат овој голем вариетет на форми, и тоа е важно затоа што влијае на нивната способност да се снабдуваат со нутриенси, да се прикрепуваат за површини, да пливаат во течности или да избегнат грабливци.[34][35]

Најголем број на бактериски видови постојат како единечни клетки, додека други може да се групираат во карактеристични форми: Neisseria се јавува во парови, Streptococcus формира ланци, Staphylococcus формира гроздовидни формации или кластери. Бактериите, исто така, може да се групираат за да формираат поголеми повеќеклеточни структури, на пр. издолжени филаменти кај цијанобактериите и актинобактериите, агрегати како кај миксобактериите, или пак комплексни хифи како кај стрептомицетите.[36] Овие повеќеклеточни структури често се формираат само при посебни услови, на пример, при недостаток на аминокиселини, миксобактериите комуницираат со околните бактерии од својот вид преку процес наречен „чувство на кворум“ (англ., quorum sensing), при што мигрираат една кон друга и се агрегираат во телца кои се со димензија од околу 500 микрометри и содржат околу 100.000 бактериски клетки.[37] Во овие повеќеклеточни телца се јавува функционална поделба меѓу одделните бактериски клетки, на пр., секоја десетта клетка од колонијата мигрира кон површината и таму се диференцира во посебна дормантна (спиечка) форма која се нарекува миксоспора, и на овој начин прават заштитен слој којшто колонијата ја штити од исушување и други влијанија на околината.[38]
Други видови на бактерии се способни да се прицврстат за површини и да формираат густи агрегати наречени биофилмови. Овие биофилмови може да варираат во дебелина од неколку микрометри па дури до половина метар, а во нив се населуваат, покрај бактериите, и разни видови на археи и еукариоти. Бактериите што живеат во биофилмови имаат сложен распоред на клетките и на вонклеточните компоненти, формирајќи секундарни структури, наречени микроколонии, помеѓу кои има мрежа од канали, кои овозможуваат подобра дифузија на хранливите материи.[39][39] Во природните средини како што се почвата, површината на камења, карпите и површината на растенијата, најголем број од бактериите се наоѓаат во површински биофилмови.[40] Биофилмовите се важни и во медицината, бидејќи овие структури често се среќаваат при хроничните бактериски инфекции или кај инфекции од имплантирани медицински помагала, затоа што бактериите што се наоѓаат во биофилмот многу потешко се уништуваат од изолираните форми.[41]
Клеточна структура
[уреди | уреди извор]
Внатрешноклеточни структури
[уреди | уреди извор]Бактериската клеточна мембрана се состои главно од фосфолипиди и белковини. Таа покрај својата заштитна улога како бариера, врши и многу важна улога во транспортните и енергетските процеси на клетката.[42] За разлика од еукариотските клетки, бактериските клетки најчесто не поседуваат мембрански органели, како што кај еукариотите се: јадро, ендоплазматски ретикулум, Голџиев систем итн.[42] Меѓутоа, некои бактерии поседуваат белковински органели во цитоплазмата кои разделуваат различни процеси од бактерискиот метаболизам,[43][44] како што се карбоксизомите.[45] Дополнително, бактериите имаат мултикомпонентен цитоскелет, кој ја контролира локализацијата на белковините и нуклеинските киселини во цитоплазмата, а исто така учествува и во процесот на клеточната делба.[46][47][48]
Многу значајни биохемиски процеси, како што е складирањето на енергија, се одвиваат како резултат на концентрациски градиенти низ мембраните, со што се создава разлика во потенцијали на ист принцип како во батериите. Недостатокот на внатрешни мембрани кај бактериите значи дека овие процеси, како што е транспортот на електрони, треба да се одвиваат низ клеточната мембрана помеѓу внатрешниот простор кој ја сочинува цитоплазмата и надворешниот простор кој ја сочинува периплазмата.[49] Меѓутоа, кај многу фотосинтетски бактерии клеточната мембрана е значително набрана и зазема голем дел од клеточниот волумен. Вака набраната мембрана гради повеќе слоеви во кои се сместени фотосинтетските комплекси, а кај зелените сулфурни бактерии може да се најдат посебни фотосинтетски телца оградени со липиди, наречени хлорозоми.[50][51]
Повеќето бактерии немаат јадро оградено со мембрана, а нивниот генетски материјал е единечен, циркуларен, бактериски хромозом, сместен во цитоплазмата во облик на телце со неправилна форма кое се нарекува нуклеоид.[52] Како и сите други живи организми, бактериите поседуваат рибозоми, кои се неопходни во процесот на биосинтеза на белковини; но структурата на бактериските рибозоми е различна од онаа кај археите и еукариотите.[53]
Некои бактерии продуцираат внатреклеточни гранули за складирање на хранливи материи, како што се гликогенот,[54] полифосфатите,[55] сулфур[56] или полихидроксиалканоати.[57] Одредени бактериски групи, како што се фотосинтетските цијанобактерии, може да продуцираат внатреклеточни гасни везикули кои им помагаат да ја регулираат својата пловност, што им овозможува да се движат нагоре или надолу во водениот столб, па така да се прилагодат на различни нивоа на светлински интензитет или концентрација на хранливи материи.[58]
Надворешноклеточни структури
[уреди | уреди извор]Над клеточната мембрана се наоѓа клеточниот ѕид. Бактерискиот клеточен ѕид е составен од пептидогликан (муреин), кој е изграден од полисахаридни ланци кои накрсно се поврзани со пептиди што содржат D-аминокиселини.[59] Бактериските клеточни ѕидови се разликуваат од растителните и габичните клеточни ѕидови, кои се изградени од целулоза и хитин, соодветно.[60] Археите, пак, за разлика од бактериите, имаат клеточен ѕид кој се состои од друг хетерополимер, наречен псевдопептидогликан. Клеточниот ѕид е неопходен за преживување на бактериите, а антибиотикот пеницилин ги убива бактериите така што инхибира еден чекор во биосинтезата на пептидогликанот.[60]
Грубо кажано, постојат два типа клеточни ѕидови кај бактериите, наречени Грам-позитивни и Грам-негативни. Имињата потекнуваат од реакција на клетките кон боењето по Грам, коешто претставува класичен метод за класификација на бактериските видови.[61]

Грам-позитивните бактерии поседуваат дебел клеточен ѕид кој содржи повеќе слоеви пептидогликан и теихоински киселини. За разлика од нив, Грам-негативните бактерии имаат релативно тенок клеточен ѕид, кој се состои од само неколку слоеви пептидогликан, врз кој имаат секундарна липидна мембрана богата со липополисахариди и липобелковини. Повеќето бактериски видови имаат Грам-негативен клеточен ѕид, а само колената Firmicutes и Actinobacteria имаат Грам-позитивен клеточен ѕид.[62] Разликите во структурата на клеточниот ѕид дава и разлика во чувствителноста спрема антибиотици, на пр. ванкомицин може да ги убие само Грам-позитивните бактерии, а не е ефикасен против Грам-негативните бактерии, како што се Haemophilus influenzae и Pseudomonas aeruginosa.[63] Некои бактерии поседуваат структура на клеточниот ѕид која не е ниту Грам-позитивна ниту Грам-негативна. Овде спаѓаат и некои клинички важни бактерии, како што се микобактериите, кои имаат клеточен ѕид изграден од дебел слој на пептидогликан, слично како кај Грам-позитивните бактерии, но поседуваат и секундарен надворешен слој од липиди.[64]
Кај многу бактерии се среќава и Ѕ-слој од ригидно аранжирани белковински молекули, кој ја покрива надворешноста на клетката.[65] Овој слој нуди физичка и хемиска заштита на клеточната површина, а делува и како бариера за макромолекуларна дифузија. Ѕ-слоевите имаат разновидни, и најчесто слабо проучени, функции, но познато е дека делуваат како фактори на вирулентност кај Campylobacter, а кај бактеријата Bacillus stearothermophilus содржат површински ензими.[66]
Камшичињата (флагели) се ригидни белковински структури, околу 20 нанометри во пречник и до 20 микрометри во должина, кои служат за моталитет (движење) на бактеријата. Енергијата за движење на камшичето се ослободува по пат на трансфер на јони преку електрохемискиот градиент којшто постои низ клеточната мембрана.[67]
Фимбриите се тенки белковински филаменти, обично 2-10 нанометри во пречник и неколку микрометри во должина. Тие се распоредени по површината на клетката и наликуваат на тенки влакненца кога се набљудуваат низ електронски микроскоп. За нив се верува дека играат улога во прицврстувањето за површини или за други клетки, и се есенцијални за вирулентноста на некои бактериски патогени.[68] Пилите (едн. лат., pilus) се клеточни структури нешто поголеми од фимбриите, кои служат за трансфер на генетски материјал помеѓу две бактериски клетки во тек на процесот на конјугација.[69] Друг вид на пили служат за движење и се нарекуваат тип IV пили.[70]
Гликокаликсот го продуцираат многу бактерии како заштитна обвивка. Тој може многу да варира во однос на структурната сложеност, од неорганизиран лигав слој составен од вонклеточни полимерни супстанци до високо структурирана капсула. Овие структури можат да ја штитат бактериската клетка од проголтување од страна на еукариотски клетки, како што се макрофагите.[71] Тие можат да служат и како антигени, и да се инволвирани во клеточното распознавање (англ., cell recognition), а можат да служат и како прицврстувач на клетката за површини и во формирањето на биофилмови.[72]
Создавањето на овие вонклеточни структури зависи од функционирањето на бактериските секреторни системи. Овие системи трансферираат белковини од цитоплазмата во периплазмата или директно во надворешноста на клетката. Познати се многу типови на секрециони системи, а овие структури честопати се есенцијални за вирулентноста на патогените, па затоа и интензивно се проучувани во медицинските науки.[73]
Ендоспори
[уреди | уреди извор]
Одредени видови на Грам-позитивни бактерии, како што се припадниците на родовите Bacillus, Clostridium, Sporohalobacter, Anaerobacter и Heliobacterium, можат да формираат високорезистентни, дормантни структури наречени ендоспори.[74] Ендоспорите се развиваат внатре во цитоплазмата, а обично само една ендоспора се развива во секоја поединечна клетка.[75]
Ендоспората во центарот содржи нуклеински материјал и рибозоми, кои се обвиткани со заштитен слој, врз кој се развива повеќеслојна ригидна обвивка составена од пептидогликан и белковини.[75] Метаболизмот внатре во ендоспората драстично се забавува, што им овозможува да преживеат екстремни физички и хемиски стресови, високи дози на ултравиолетово зрачење, гама-зрачење, високи и ниски температури, висок притисок, сушење, дезифициенси итн.[76] Во ваква дормантна состојба бактериите можат да преживеат милиони години,[77][78] а исто така е покажано дека можат да преживеат изложување на вакуум и зрачење во вселената.[79] Ендоспорите на некои бактериски видови можат да бидат патогени, на пр., заразувањето со антракс настанува со вдишување на ендоспорите на Bacillus anthracis, а контаминацијата на длабоки убодни рани со ендоспори на Clostridium tetani предизвикува тетанус.[80]
Метаболизам
[уреди | уреди извор]Бактерискиот свет е познат по тоа што изобилува со различни типови на метаболизам.[81] Метаболистичките особини на бактериите во склоп на една група традиционално биле користени за да се одреди нивната таксономија, што подоцна се покажало дека не се совпаѓа потполно со најновите генетски класификации.[82] Бактерискиот метаболизам е класифициран во нутрициони групи врз основа на три основни критериуми: типот на енергија која се користи за раст, изворот на јаглерод и дарителите на електрони. Дополнителен критериум за дишните бактерии се акцепторите на електрони, според кој бактериите се делат на анаеробни и аеробни.[83]
Јаглеродниот метаболизам кај бактериите е или автотрофен (извор на јаглерод е јаглерод диоксидот) или хетеротрофен (извор на јаглерод се органските соединенија). Типични автотрофни бактерии се цијанобактериите, зелените сулфурни бактерии и пурпурните бактерии, но, исто така, тука се вбројуваат и хемолитотрофните видови, како што се нитрификационите и сулфур-оксидирачките бактерии.[84] Енергетскиот метаболизам кај бактериите е заснован или на фототрофија (користење на светлинска енергија преку процесот на фотосинтеза) или на хемотрофија (користење на хемиски супстанци како извор на енергија, преку нивната оксидација со кислород или други акцептори на електрони).
Понатамошната поделба е на литотрофи, кои користат неоргански дарители на електрони, и органотрофи, кои користат органски дарители на електрони. Хемотрофните бактерии ги користат дарителите на електрони во процесите на складирање на енергија (аеробно/анаеробно дишење или ферментација) и во биосинтетските реакции, додека фототрофните бактерии ги користат само во биосинтетските реакции. Дишните бактерии користат разни соединенија како извор на енергија, со тоа што одземаат електрони од редуцираните супстрати, а потоа ги трансферираат овие електрони на терминален акцептор на електрони во реакција на оксидо-редукција. Ваквите реакции ослободуваат големо количество на енергија, кое може да биде искористено во синтезата на аденозин трифосфат (ATP). Кај аеробните организми, терминалниот акцептор на електрони е кислородот, додека кај анаеробните организми оваа улога можат да ја вршат различни неоргански соединенија, како што се сулфатот, нитратот, јаглерод диоксидот итн. Ова води до еколошки значајни процеси, како што се редукцијата на сулфатите, денитрификацијата и ацетогенезата, соодветно.[85][86][87][88]

Во услови на недостаток на акцептори на електрони, друг облик на метаболизам кај хемотрофите е ферментацијата. При ферментација, електроните одземени од редуцираните супстрати се трансферираат на оксидирани меѓупроизводи (интермедиери), со што се создаваат редуцирани ферментациони производи (на пр., млечна киселина, етанол, водород, бутаноинска киселина). Ферментацијата е возможна затоа што енергетската содржина на супстратите е поголема од онаа на производите, што му овозможува на организмот да синтетизира ATP во текот на овој процес.[89] Факултативните анаероби, пак, имаат способност да во зависност од условите на средината користат или ферментација или дишење за своите енергетски потреби.
Литотрофните бактерии се способни да користат неоргански соединенија како извор на енергија. Најчести неоргански дарители на електрони се водородот, јаглерод моноксидот, амонијакот (што доведува до нитрификација),[90] двовалентни железни јони (Fe2+) и други редуцирани метални јони,[91] како и некои редуцирани сулфурни соединенија. Во некои услови, гасот метан може да се користи од страна на метанотрофните бактерии како извор на електрони и како супстрат за јаглеродниот анаболизам.[92] Кај аеробната фототрофија и хемолитотрофија, кислородот се користи како терминален акцептор на електрони, додека при анаеробни услови можат да се користат и други неоргански соединенија. Повеќето литотрофни бактерии се автотрофни, додека органотрофните бактерии по правило се хетеротрофни.
Освен тоа што некои бактерии се способни да го користат јаглерод диоксидот од атмосферата како извор на јаглерод, други видови на бактерии се способни да го врзуваат азотот од атмосферата со помош на ензим наречен нитрогеназа.[93]
Без разлика кој тип на метаболизам го употребуваат, мнозинството од бактерискиот свет е способен да ги прима суровините од околината само во форма на релативно мали молекули кои навлегуваат внатре во клетката или со дифузија или преку специјални белковински канали во клеточната мембрана. Исклучок се планктомицетите, единственото колено на бактерии кои поседуваат мембрани околу јадрениот материјал. Неодамна било покажано дека видот Gemmata obscuriglobus е способен да внесува големи молекули во клетката преку процес кој наликува на ендоцитоза, слично како кај еукариотските клетки.[30][94]
Раст и репродукција
[уреди | уреди извор]Бактериите растат до одредена големина, а потоа се размножуваат по пат на бинарна (проста) делба, што е форма на бесполово размножување.[95] При оптимални услови бактериите може да растат и да се делат исклучително брзо, а бактериските популации може да се дуплираат во големина на секои 9,8 минути.[96] При оваа проста делба се создаваат две идентични клетки - клонови. Некои бактерии кои се размножуваат бесполово формираат комплексни репродуктивни структури кои помагаат за дисперзирање на новосоздадените клетки-ќерки. Примери се: плодоносното тело кај Myxobacteria и воздушните хифи кај Streptomyces. Начин на бесполова репродукција е и пупењето, кога кај родителската клетка се формира израсток (пупка) кој потоа се откинува и ја дава клетката-ќерка.

Во лабораторија, бактериите главно се одгледуваат во цврсти или течни медиуми. Цврстите медиуми за раст ги вклучуваат агарните плочи и се користат да се изолира чиста култура на одреден бактериски вид. Течните медиуми за раст се користат кога е потребно мерење на растот или кога се потребни поголеми волумени на клетки. Растот во течни медиуми кои се мешаат создава рамномерна суспензија на клетки, што овозможува културите лесно да се раздвојат и трансферираат на друга подлога. Сепак, изолацијата само на еден бактериски вид од течниот медиум е тешка. Употребата на селективни медиуми, во кои има додаток или недостаток на специфични хранливи состојки или антибиотици, може да помогне да се идентификуваат специфични бактерии.[97]
Повеќето лабораториски техники за одгледување на бактериите користат високи концентрации на хранливи материи за да се добијат големи количини на клетки брзо и евтино. Меѓутоа, во природните средини хранливите материи се ограничени, што значи дека бактериите не можат бесконечно да се размножуваат. Овој лимит на нутриенси (хранливи состојки) довел до еволуција на различни стратегии за раст (види: r/K селекциона теорија). Некои бактерии можат да растат исклучително брзо при достапност на нутриенси, како што е, на пример, цветењето на цијанобактериите на езерски површини во летните месеци. Други бактерии развиле адаптација на сурови средини, како што се средини со Streptomyces видови кои излачуваат голем број на разни антибиотици за да го инхибираат развојот на околните компетитивни бактерии.[98] Во природата, најголем број бактерии живеат во заедници наречени биофилмови, кои овозможуваат стабилно снабдување со нутриенси и заштита од факторите на околината.[40] Меѓусебните односи внатре во заедницата се есенцијални за растот и размножувањето на специфични организми или групи на организми (тип на симбиоза што се нарекува синтрофија).[99]
Бактерискиот раст се одвива во четири фази. Кога бактериска популација првпат стапува во контакт со високохранлива средина, потребно е да помине некое време во кое клетките се адаптираат на новата средина. Првата фаза од растот е наречена лаг-фаза, што претставува период на бавен раст кога клетките се адаптираат на новата средина и се подготвуваат за раст. Во лаг-фазата е забележана зголемена брзина на метаболизмот, а клетката продуцира разни белковини и ензими неопходни за следната фаза на брз раст.[100] Втората фаза на раст е лог-фаза (логаритамска или експоненцијална фаза), која се одликува со експоненцијален раст. Брзината со којашто клетките растат во текот на оваа фаза е позната како брзина на раст (k), додека времето што им е потребно да се дуплираат во број се нарекува генерациско време (g). Во текот на лог-фазата, нутриенсите се метаболизираат со максимална брзина, сѐ додека еден од нутриенсите не почне да се троши и со тоа да го лимитира понатамошниот раст. Третата фаза на растот се вика стационарна фаза, и е предизвикана од потрошување на нутриенсите. Клетките почнуваат да ја редуцираат метаболистичката активност и да разградуваат некои неесенцијални клеточни белковини. Стационарната фаза е транзиција од брз раст во состојба на одговор кон стрес, како што е зголемена експресија на гени инволвирани во репарација на ДНК, гени на антиоксидативниот метаболизам и гени за транспорт на нутриенси.[101] Последната фаза е фаза на смрт, кога бактериите целосно ги исцрпуваат нутриенсите, по што умираат.
Геноми и хромозоми
[уреди | уреди извор]До септември 2016 година, секвенционирани се геномите на илјадници бактериски видови, од кои околу 9.000 се комплетирани, а повеќе од 42.000 се во драфт-форма.[102]
Повеќето бактерии имаат еден циркуларен хромозом, кој може да варира во големина од само 160.000 базни парови (bp), како кај ендосимбиотската бактерија Candidatus Carsonella ruddii,[103] па сѐ до 12.200.000 базни парови (12,2 Mbp) како кај почвената бактерија Sorangium cellulosum.[104] Гените во бактерискиот хромозом најчесто се единечен континуиран сегмент од ДНК молекулата, и иако постојат неколку различни типови на интрони кај бактериите, тие се многу поретки отколку кај еукариотите.[105]
Повеќе од еден хромозом се среќава кај родот Vibrio, на пр., Vibrio cholerae, причинителот на колера, и V. parahaemolyticus, имаат по два хромозома.[106] Впрочем, 10% од сите секвенционирани бактериски геноми имаат два или повеќе хромозоми.[106]
Некои бактерии, како на пр., Borrelia burgdorferi, причинителот на лајмска болест, содржат еден линеарен хромозом и неколку линеарни или циркуларни плазмиди.[107][108] Хромозомот на Streptomyces lividans е исто така линеарен, а постои доказ дека ова важи за повеќето видови на овој род.[109]
Плазмидите се мали екстрахромозомски ДНК молекули кои содржат гени за: ензими кои ги разградуваат невообичаените органски супстрати, ензими за стекнување резистенција кон антибиотици, ензими за синтеза на токсини кои ги убиваат другите бактерии, или ензими за фактори на вирулентност.[110] Плазмидите се реплицираат независно од хромозомите и често кодираат т.н. партиционен систем кој осигурува дека секоја клетка-ќерка ќе добие копија од плазмидот во текот на клеточната делба. Партиционите системи, исто така, можат да бидат кодирани од хромозомската ДНК. Плазмидите кои се наоѓаат во голем број на копии во една клетка, најчесто, немаат партициони системи, бидејќи шансата да се распоредат во двете клетки-ќерки е голема.[111]
Генетика
[уреди | уреди извор]Бактериите како бесполови организми се клонали, што значи дека наследуваат идентична копија од гените на родителот. Поради ова, еволуцијата кај бактериите настанува како резултат на селекција на одредени мутации, или пак како резултат на промени во нивниот генетски материјал стекнати со генетска рекомбинација. Мутациите настануваат како резултат на грешки во текот на репликацијата на ДНК или поради изложување на мутагени фактори. Степенот на мутирање широко варира кај разните видови бактерии, а дури и меѓу различните клонови на една иста бактерија.[112] Генетските промени во бактерискиот геном настануваат или со случајни мутации или со „мутации индуцирани од стрес“, каде што гените инволвирани во одреден процес кој го лимитира растот имаат покачен степен на мутирање.[113]
Трансфер на ДНК
[уреди | уреди извор]Некои бактерии се способни да трансферираат генетски материјал помеѓу клетките, што може да се одвива на три различни начини. Прво, бактеријата може да прими егзогена ДНК од својата околина, што претставува процес наречен трансформација. Трансферот на гени може да се случи и преку процес наречен трансдукција, што се случува кога бактериофаг ја интегрира својата ДНК во хромозомот на бактеријата-домаќин. Третиот метод за трансфер на гени се нарекува конјугација и настанува кога сегменти од ДНК се трансферираат со директен контакт на две бактериски клетки.

Трансдукција на бактериски гени, од страна на бактериофаги, настанува како резултат на грешки во тек на интеграцијата на вирусните честики во цитоплазмата. Процесот на конјугација, како што се јавува кај E. coli, е детерминиран од плазмидски гени, кои ја кодираат машинеријата којашто е потребна за да се трансферира новата копија од плазмидската ДНК од бактеријата-дарител на бактеријата-акцептор. Понекогаш се случува конјугативниот плазмид да се интегрира во хромозомот на бактеријата-домаќин и, на тој начин, да се трансферира дел од хромозомската ДНК на друга бактерија.
За разлика од трансдукцијата и конјугацијата, трансформацијата е зависна од мноштво производи на бактериските гени, кои стапуваат во специфична интеракција за да го изведат овој сложен процес.[114] За да може бактеријата да врзе, да прими и да рекомбинира дарителска ДНК во својот хромозом, најпрво мора да влезе во специјална физиолошка состојба наречена природна компетентност. Кај Bacillus subtilis потребни се околу 40 гени за да се развие компетентност.[115] Должината на ДНК молекулата, која се трансферира во тек на трансформацијата на Bacillus subtilis, може да биде од третина хромозом, па сѐ до цел хромозом.[116] Се претпоставува дека трансформацијата е честа во бактерискиот свет и до сега се идентификувани најмалку 60 видови кои ја имаат природната способност да станат компетентни за трансформација.[117] Развојот на компетентноста во природата често е поврзан со условите на околината кои предизвикуваат стрес, и, се чини, дека таа е начин на адаптација со кој се олеснува репарацијата на оштетената ДНК во клетките-реципиенти.
При обични услови, трансдукцијата, конјугацијата и трансформацијата вклучуваат трансфер на ДНК меѓу бактерии од ист вид, но во одредени случаи трансферот може да се случи и меѓу единки од различни видови.[118] Во овие случаи, примањето на гени од други бактерии или од околината е наречен хоризонтален пренос (трансфер) на гени, и во одредени природни услови може да биде многу често застапен.[119] Трансферот на гени е особено значаен за стекнување на резистентност кон антибиотици, бидејќи овозможува брз трансфер на гени кои нудат резистентност помеѓу различни видови на патогени бактерии.[120]
Бактериофаги
[уреди | уреди извор]Бактериофагите се вируси кои ги инфицираат бактериите. Постојат многу типови на бактериофаги; некои едноставно ја инфицираат и лизираат бактеријата-домаќин, додека други се инкорпорираат во бактерискиот хромозом. Бактериофагот може да содржи гени кои го менуваат фенотипот на домаќинот; на пример, во еволуцијата на Escherichia coli O157:H7 и Clostridium botulinum, гените за токсини на интегрираниот бактериофаг ги имаат преобразено безопасните предци на овие бактерии во смртоносни патогени.[121] Бактерискиот одбранбен одговор кон инфекција со бактериофаг се одвива преку рестрикциони модифицирачки системи, кои ја деградираат туѓата ДНК,[122] и друг систем кој ги користи CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) секвенците за да зачува фрагменти од геномот на бактериофагот, со кој бактеријата веќе дошла во контакт порано, што им овозможува да ја блокираат репликацијата на вирусот со помош на РНК интерференција.[123][124] На овој начин, CRISPR системот ѝ дава на бактеријата здобиен имунитет кон инфекцијата.
Однесување
[уреди | уреди извор]Движење
[уреди | уреди извор]
Многу видови на бактерии се способни да се движат користејќи различни механизми: камшичињата се користат за пливање во течност; бактериското лизгање и трепкачкиот моталитет служат за движење по површини; а промените во пловноста овозможуваат вертикално движење во течност.[125]
Бактериите кои пливаат, најчесто поминуваат растојание 10 пати поголемо од нивната големина во секунда, а само неколку видови можат да поминат растојание 100 пати поголемо од нивната големина во секунда. Кај бактериското лизгање и моталитетот со трепкање, бактеријата ги употребува своите тип IV пили како прицврстувач, издолжувајќи ги, усидрувајќи ги и на крај повлекувајќи ги со значителна сила, поголема од 80 pN (пикоњутни).[126]
Камшичињата (флагели) се семиригидни, цилиндрични структури кои се ротираат на начин сличен на пропелер на брод. Бактериските видови се разликуваат по бројот и распоредот на флагелите на нивната површина; некои имаат едно камшиче (монотрихни); други имаат камшичиња поставени на двата краја од клетката (амфитрихни); некои имаат снопови од флагели на двата пола од клетката (лофотрихни); додека некои имаат флагели распоредени по целата површина на клетката (перитрихни). Бактериското камшиче е најдобро проучената структура за моталитет во биологијата, а изградено е од околу 20 различни белковини, со околу 30 други белковини неопходни за негова регулација и изградба.[125] Камшичето е вртечка структура која ја покренува реверзибилен „мотор“, кој се наоѓа на базата од камшичето и користи електрохемиски градиент преку мембраната како извор на енергија.[127] На овој начин моторот го покренува движењето на филаментот, којшто игра улога на пропелер.
Најголем број на бактерии имаат два различни начина на движење; праволиниско движење (пливање) и свртување. Свртувањето им овозможува да се реориентираат, што го прави нивното севкупно движење тридимензионално случајно шетање.[128] Флагелата кај спирохетите е сместена меѓу двете мембрани во периплазматскиот простор. Тие имаат уникатна форма на спирала, а движењето наликува на вртењето на сврдло.[125] Мобилните бактерии се движат кон или подалеку од одредени стимуланси, а овие однесувања збирно се нарекуваат такси: хемотакса, фототакса, магнетотакса и енергетска-такса.[129]
Некои видови на Listeria и Shigella се движат внатре во клетката на домаќинот така што го узурпираат нејзиниот цитоскелет, кој нормално се користи за движење на органелите на домаќинот. Со тоа што поттикнуваат полимеризација на актин на едниот пол од сопствената клетка, тие можат да формираат опавче со кое се движат низ цитоплазмата на домаќинот.[130]
Секреција
[уреди | уреди извор]Бактериите често секретираат различни супстанци во околината со цел да ја модифицираат во нивна полза. Секретираните соединенија често се белковини, кои може да служат како ензими за дигестија на некои хранливи материи во околината.
Повеќеклеточност
[уреди | уреди извор]Бактериите често функционираат како повеќеклеточни (мултицелуларни) агрегати наречени биофилмови, во кои тие разменуваат молекуларни сигнали за меѓуклеточна комуникација, што води до координирано повеќеклеточно однесување.[131][132] Погодностите на заедницата при мултицелуларната соработка вклучуваат: поделба на трудот, пристап до ресурси кои не можат ефикасно да ги искористат поединечните клетки, колективна одбрана од антагонисти и оптимизација на преживувањето на популацијата со диференцијација во различни типови на клетки. На пример, бактериите во биофилмовите можат да имаат повеќе од 500 пати зголемена резистентност кон антибактериски агенси во споредба со поединечните (планктонски) бактерии од истиот вид.
Еден од типовите на меѓуклеточната комуникација со молекуларни сигнали е т.н. „чувство на кворум“ (англ., quorum sensing), кој има за цел да одреди дали густината на локалната популација е доволно висока за да биде продуктивно да се инвестира во процеси кои се успешни само кога голем број на слични организми се однесуваат слично, како што е, на пр., излачувањето на дигестивни ензими или емитирање на светлина. Чувството на кворум им овозможува на бактериите да ја координираат експресијата на гени, што им овозможува да продуцираат, ослободуваат и детектираат автоиндуцирачки молекули или феромони, кои се акумулираат со растот на бактериската популација.[133]
Биолуминисценција
[уреди | уреди извор]Неколку бактериски видови имаат хемиски системи за создавање на светлина. Оваа особина се нарекува биолуминисценција и се среќава кај бактерии кои живеат во асоцијација со риби, а светлината веројатно служи за привлекување на други животни.
Класификација и идентификација
[уреди | уреди извор]
Класификацијата има за цел да го опише диверзитетот на бактериските видови со нивно именување и организирање во групи врз основа на сличности. Бактериите може да се класифицираат на основа на нивната клеточна структура, метаболизмот или разликите во клеточните компоненти, како што се ДНК, РНК, масни киселини, пигменти, антигени, хинони итн. Додека овие критериуми овозможиле идентификација и класификација на типови на бактерии, не било јасно дали овие разлики претставуваат варијација помеѓу различни видови или помеѓу подгрупи кои припаѓаат на истиот вид. Оваа несигурност се должела на недостатокот на својствени структури кај повеќето бактерии, како и на хоризонталниот трансфер на гени помеѓу несродните видови.[134] Како резултат на хоризонталниот трансфер на гени, блиску сродните бактерии можат да имаат многу различна морфологија и метаболизам. За да се надмине оваа несигурност, модерните бактериски класификации се засноваат на молекуларна систематика, за која се употребуваат генетски техники, како што се одредувањето на односот гванин:цитозин (G:C), геном-геном хибридизација, како и секвенционирање на гените кои не подлегнале на екстензивен хоризонтален трансфер на гени, како што е генот на рибозомната РНК.[135] Класификацијата на бактериите се официјализира со објавување во Меѓународното списание за систематска бактериологија (англ., International Journal of Systematic Bacteriology) и Бергејовиот прирачник за систематска бактериологија (англ., Bergey's Manual of Systematic Bacteriology). Меѓународниот комитет за систематика на прокариотите (англ., International Committee on Systematics of Prokaryotes - ICSP) ги одржува интернационалните правила за именување на бактериите и таксономските категории и за нивно рангирање во Меѓународниот кодекс за номенклатура на бактериите (англ., International Code of Nomenclature of Bacteria).
Терминот „бактерии“ традиционално ги опфаќал сите микроскопски едноклеточни прокариоти. Меѓутоа, молекуларните систематики покажале дека прокариотскиот свет се состои од два одделни домени, првично наречени еубактерии и архебактерии (денес се користат називите бактерии и археи), кои еволуирале независно од еден заеднички предок.[16] Моментално најупотребуваниот класификационен систем во биологијата е т.н. систем на трите домени, според кој бактериите, археите и еукариотите се разгледуваат како три основни групи на живи организми на планетата.[136] Како резултат на релативно скорешното воведување на молекуларната систематика и постојаниот раст во бројот на секвенционираните геноми кои се достапни, бактериската класификација сѐ уште останува поле на истражување кое е променливо и се шири.[137]
Идентификацијата на бактериите во лабораторија е од посебно значење за медицината бидејќи одредувањето на точната терапија зависи од бактерискиот вид кој е причинител на инфекцијата. Боењето според Грам, откриено во 1884 година од страна на Ханс Кристијан Грам, ги карактеризира бактериите според структурата на нивниот клеточен ѕид. Дебелите слоеви од пептидогликан кај Грам-позитивниот клеточен ѕид се обојуваат пурпурно, додека тенките слоеви од пептидогликан кај Грам-негативниот клеточен ѕид се обојуваат розово. Со комбинација на морфологијата и Грам-обојувањето, повеќето бактерии може да се класифицираат во 4 групи: Грам-позитивни коки, Грам-позитивни бацили, Грам-негативни коки и Грам-негативни бацили. Некои бактерии најдобро се идентификуваат со обојувања различни од она на Грам, особено микобактериите и Nocardia, кои се бојат по Цил-Нилсен или слични обојувања.[138] Некои бактерии може да се идентификуваат само врз основа на нивниот раст во специјални медиуми или со употреба на техники како што е серологијата.
Техниките со култура се дизајнирани да го поттикнат растот на одредени бактерии, додека се забавува растот на други бактерии во примерокот, што помага за идентификација. Често овие техники се дизајнирани за специфични примероци, како на пр., примерок од плунка ќе биде третиран за идентификација на причинителот на пневмонија, додека примероци од измет се одгледуваат на селективни медиуми за да се идентифицира причинителот на дијареја, додека во исто време се спречува растот на непатогените бактерии. Примероците кои треба да бидат стерилни, како што се крвта, ликворот, урината, се одгледуваат во посебни услови дизајнирани да се овозможи раст на сите можни микроорганизми.[139] Штом ќе се изолира патоген микроорганизам, се пристапува кон негова идентификација врз основа на морфологијата, одлики во растот, обојувањето, хемолитичките особини итн.
Во идентификацијата на бактериите, сѐ почесто се користат молекуларните методи. Дијагностиката заснована на ДНК методи, како што е полимераза верижната реакција (PCR) стануваат сѐ попопуларни, главно како резултат на нивната специфичност и брзина.[140] Овие методи овозможуваат детекција и идентификација на вијабилни (живи) бактерии кои не можат да се одгледуваат, кои се метаболистичко активни, но не се делат.[141] Меѓутоа, дури и со употреба на овие современи методи, вкупниот број на бактериски видови е непознат и не може со сигурност да се одреди. Според денешните класификации има нешто помалку од 9.300 познати видови на прокариоти, вклучувајќи ги бактериите и археите,[142] но проценките за вистинскиот број на бактерискиот диверзитет се движи во рамките на 107 - 109 бактериски видови, иако постојат мислења дека и овие цифри го потценуваат вистинскиот диверзитет на бактерискиот свет.[143][144]
Заемодејства со други организми
[уреди | уреди извор]Бактериите можат да формираат комплексни асоцијации со други организми. Тие се делат на: мутуализам, коменсализам и паразитизам. Како резултат на нивната мала величина, коменсалните бактерии се убиквитарни и растат на растенијата и животните на ист начин на кој би растеле и на која било друга површина. Меѓутоа, нивниот раст може да се зголеми при топлина и потење, и големите популации на овие бактерии кај луѓето се причина за специфичен мирис на телото.
Грабливци
[уреди | уреди извор]Некои видови на бактерии убиваат и консумираат други микроорганизми, а тие се наречени грабливи бактерии.[145] Пример за оваа група е Myxococcus xanthus, која формира роеви од клетки кои убиваат и дигестираат кој било друг вид на бактериски клетки.[146] Има грабливци кои се прицврстуваат за нивниот плен со цел да го дигестираат и да му ги апсорбираат нутриенсите, како што е бактеријата Vampirovibrio chlorellavorus, додека пак други ја инвадираат клетката и се делат во нејзината цитоплазма, како што прави Daptobacter.[147] Овие грабливи бактерии се смета дека еволуирале од детривори, кои консумираат мртви микроорганизми, преку адаптации кои им овозможиле да фаќаат и убиваат живи микроорганизми.
Мутуалисти
[уреди | уреди извор]
Пример за мутуалистичко поврзување е т.н. водороден трансфер, кој се одвива меѓу кластери од анаеробни бактерии, кои консумираат органски киселини и продуцираат водород, и метаногени археи, кои го консумираат водородот.[148] Бактериите во овој тип на асоцијација сами за себе не можат да ги консумираат органските киселини, бидејќи овој процес ослободува големо количество на водород, кој ослободен во околината го забавува нивниот раст. Само кога се наоѓаат во асоцијација со водород-консумирачките археи, концентрацијата на водородот би била доволно ниска за да им овозможи нормален раст.
Во почвата, бактериите кои се наоѓаат во ризосферата (зона која вклучува површина на корен и почвени зрнца кои остануваат залепени за него после благо тресење) вршат фиксација на азот, претворајќи го атмосферскиот азот во разни азотни соединенија.[149] На овој начин овие т.н. азотофиксирачки бактерии им обезбедуваат на растенијата достапна форма на азот. Многу други бактериски видови градат симбиотски врски со животните, вклучувајќи го и човекот. Повеќе од илјада бактериски видови ја чинат нормалната човекова микрофлора на цревата, која придонесува за имунитет, синтеза на есенцијални витамини (витамин К, фолна киселина, биотин), претворање на шеќери во млечна киселина, како и ферментација на несварливи комплексни јаглехидрати.[150][151] Присуството на цревната микрофлора, исто така, го инхибира растот на потенцијално патогените бактерии (најчесто преку компетитивна ексклузија) и овие корисни бактерии се тие кои се продаваат како пробиотски прехранбени додатоци.[152]
Патогени
[уреди | уреди извор]Ако бактериите формираат паразитска асоцијација со други организми тие се сметаат за патогени. Тие се причинители на голем број болести кај човекот како што се: туберкулоза, тетанус, дифтерија, тифус, сифилис, колера, лепра, лајмска болест итн. Патогената инфективна причина кај некои болести може да се открие години по појавата на болеста, како што е случајот со Helicobacter pylori, предизвикувачот на пептички чир. Бактериските болести се значајни и во земјоделството, каде бактериите се причинители на разни растителни заболувања, а исто така и на заболувања кај домашните животни.
.jpg?lang=mk)
Секој вид патогена бактерија има карактеристичен спектар на заемодејства со својот домаќин. Некои бактерии, како што се стафилококите и стрептококите, можат да предизвикаат: кожни инфекции, дишни инфекции, менингит, сепса итн. Сепак, овие видови можат да бидат дел од нормалната човекова флора, и тогаш живеат на кожата или во носот без да предизвикуваат болест. Други бактерии безусловно предизвикуваат болести кај човекот, како, на пр., рикецијата која е облигатен внатреклеточен паразит, способна да расте и да се размножува само во клетката на домаќинот. Други облигатно внатреклеточни паразити се хламидиите, кои можат да предизвикаат пневмонија, инфекции на мочниот и половиот систем, а може да се инволвирани и во коронарна артериска болест.[153] Видовите како: Pseudomonas aeruginosa, Burkholderia cenocepacia и Mycobacterium avium, се патогени опортунисти и причинуваат болест главно кај луѓе кои имаат имуносупресија или цистична фиброза.[154][155]
Бактериските инфекции се третираат со антибиотици, кои се класифицираат на бактерицидни (ги убиваат бактериите) и бактериостатски (го забавуваат нивниот раст). Постојат многу типови на антибиотици, а секоја класа инхибира одреден клеточен процес присутен во бактериската клетка, а кој го нема во клетката на домаќинот. Еден од примерите за тоа како антибиотиците предизвикуваат ваква селективна токсичност е хлорамфениколот, кој ја инхибира функцијата на бактерискиот рибозом, но не може да го инхибира структурно различниот еукариотски рибозом.[156] Покрај во хуманата медицина, антибиотиците масовно се употребуваат и во т.н. интензивно сточарство, каде имаат улога да го забрзаат растот на животните, што може да има негативен ефект во смисла на брзо развивање на антибиотска резистентност кај бактериските популации.[157] Инфекциите од бактерии можат да бидат спречени со примена на антисептички мерки, како што е стерилизација на дел од кожата со алкохол пред боцкање со игла. Хируршките и стоматолошките инструменти задолжително се стерилизираат за да се спречи нивната контаминација со бактерии. Дезинфекционите средства се користат да се убијат бактериите и другите патогени микроорганизми за да се спречи контаминацијата и да се редуцира ризикот од инфекција.
Значење во индустријата и технологијата
[уреди | уреди извор]Илјадници години наназад луѓето ги користеле бактериите во производството на прехранбени продукти. Бактериите на млечно-киселинското вриење, како што се Lactobacillus и Lactococcus, заедно со разни видови на габи, сѐ уште се користат во подготовката на ферментирани производи, како на пример сирењето, јогуртот, оцетот, виното, соја сосот итн.[158]
Способноста на бактериите да разградуваат разни видови на органски соединенија се искористува во процесот на обработка на отпади и во биоремедијацијата. Бактериите кои се способни да ги дигестираат нафтените јаглеводороди се користат за чистење на истурена нафта,[159] а има видови на бактерии кои се користат за биоремедијација на токсични индустриски отпади.[160] Во хемиската индустрија, бактериите се важни во производството на енантиомерно-чисти хемикалии, кои се применуваат како лекови или како агрохемикалии.[161] Во земјоделството, бактериите се користат како замена за пестициди во т.н. биолошка контрола на штетници. Овде најчесто се користи Bacillus thuringiensis, Грам-позитивна бактерија која е честа во почвите. Еден подвид на оваа бактерија се користи како специфичен инсектицид за некои групи на инсекти.[162] Поради својата специфичност, овие пестициди не се штетни за околината, за дивиот свет и за инсектите-опрашувачи.[163]
Поради способноста да растат брзо и лесно, бактериите се најкористени организми во молекуларната биологија, генетиката и биохемијата. Со мутирање на нивната ДНК и испитување на резултирачките фенотипови, научниците ја одредуваат функцијата на гените, ензимите и метаболистичките патишта во бактериите, кои податоци можат да се користат и во изучувањето на покомплексните организми.[164] Разбирањето на бактерискиот метаболизам овозможува нивна примена во биотехнологијата, каде подлегнуваат на биоинженерство за производство на терапевтски белковини, како што се инсулинот, факторите на раст и антителата.[165][166]
Историја на бактериологијата
[уреди | уреди извор]
Развојот на микробиологијата започнува со откривањето на микроскопот од страна на холандскиот трговец Антони ван Левенхук.[167] Тој успеал да направи микроскоп со помош на комбинација на оптички леќи кои зголемувале околу 300 пати. Гледајќи низ микроскопот видел мноштво на подвижни суштества во капка од барска вода, капка од сопствената плунка и други материјали, кои ги нарекол „animalcula“. Ова откритие го презентирал во 1676 година на Кралското друштво во Лондон (англ., Royal Society of London).[168][169][170] За жал, на ова откритие во тогашното општество не му било придадено големо значење.
Германскиот природонаучник, Кристијан Готфрид Еренберг, прв го вовел терминот „бактерија“ во 1828 година, со кој тој ги именувал сите стапчести бактерии кои не формираат ендоспори, за разлика од терминот „бацили“, со кој ги именувал стапчестите бактерии кои формираат ендоспори.[171]
Најголем придонес за развојот на микробиологијата и имунологијата дал францускиот научник и хемичар, Луј Пастер. Во 1859 година, тој прв демонстрирал дека процесот на ферментација се должи на метаболистичките активности на микроорганизми, а не на спонтани хемиски реакции како што дотогаш се верувало.[172] Поаѓајќи од овие резултати, тој прв дошол до помислата дека микроорганизмите се тие кои предизвикуваат заболувања кај луѓето и животните на сличен начин како што предизвикуваат расипување на прехранбените производи со ферментација. Негови пронајдоци кои до ден денес се користат во микробиологијата се: микробиолошката еза, вештачките течни хранливи подлоги, стерилизацијата на лабораториските инструменти (на сува топлина и на пареа под притисок), пастеризацијата (стерилизација на течностите осетливи на високи температури) итн.
Усовршувањето на бактериолошките техники е заслуга на германскиот лекар и микробиолог Роберт Кох, кој поради тоа го нарекуваат „татко на бактериолошката техника“. Тој ги поставил познатите „постулати на Кох“, кои и денес се користат во медицината.[173] За откривањето на бактерискиот причинител на туберкулозата, во 1905 година тој ја добил Нобеловата награда за физиологија или медицина.[174] Во истиот период, данскиот бактериолог Ханс Кристијан Грам го пронашол боењето на бактериите наречено според него,[175] со што уште повеќе се усовршила бактериолошката техника. Овој период, во кој се откриени голем број на причинители на заразни болести, познат е како „златниот век на бактериологијата“.
Современата хемиотерапија започнува со трудовите на Пол Ерлих во почетокот на 20 век. Тој работел на полето на имунологијата и заедно со белгискиот микробиолог Жил Борде биле застапници на теоријата на хуморалниот имунитет.[176] Во истиот период на ова поле работел и рускиот биолог и патолог Иљја Мечников, кој ја поддржувал својата теорија на клеточен имунитет.[177]
Најголемиот исчекор во полето на еволуционата микробиологија го направил американскиот биофизичар Карл Воуз во 1977 година, кој користејќи ги новите молекуларни методи во филогенетиката и таксономијата, засновани на секвенционирање на 16S рРНК молекулата, за првпат ги класифицирал археите како посебен домен на животот. Со тоа тој ги поставува темелите на денешниот систем на трите домени на животот за класификација на живите организми.[16]
Галерија
[уреди | уреди извор]-
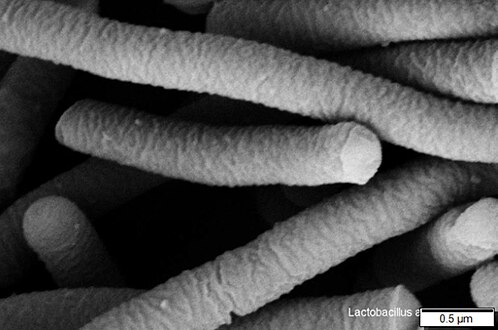 Lactobacillus acidophilus (Firmicutes)
Lactobacillus acidophilus (Firmicutes) -
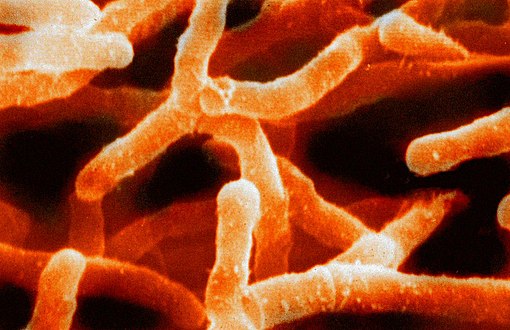
 Thermus aquaticus (Deinococcus-Thermus)
Thermus aquaticus (Deinococcus-Thermus) -
 Nostoc commune (Cyanobacteria)
Nostoc commune (Cyanobacteria) -
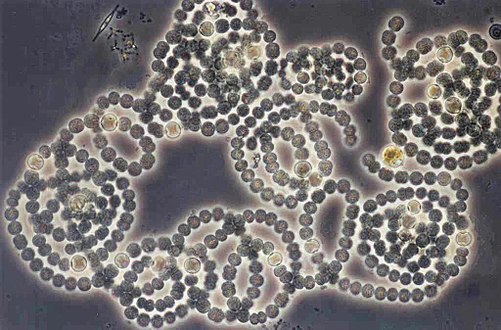 Anabaena circinalis (Cyanobacteria)
Anabaena circinalis (Cyanobacteria) -
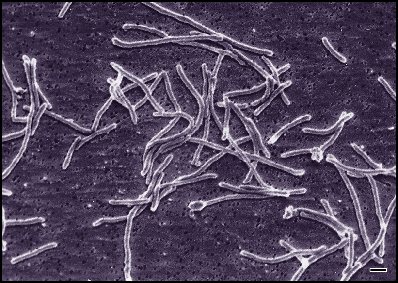
 Actinomyces israelii (Actinobacteria)
Actinomyces israelii (Actinobacteria) -
 Gemmata obscuriglobus (Planctomycetes)
Gemmata obscuriglobus (Planctomycetes) -
 Borrelia burgdorferi (Spirochaetes)
Borrelia burgdorferi (Spirochaetes) -
 Bacteroides fragilis (Bacteroidetes)
Bacteroides fragilis (Bacteroidetes) -
 Venenivibrio stagnispumantis (Aquificae)
Venenivibrio stagnispumantis (Aquificae) -
 Helicobacter pylori (Proteobacteria)
Helicobacter pylori (Proteobacteria) -
 Vibrio cholerae (Proteobacteria)
Vibrio cholerae (Proteobacteria) -
 Neisseria gonorrhoeae (Proteobacteria)
Neisseria gonorrhoeae (Proteobacteria)






.jpg?lang=mk)





Поврзано
[уреди | уреди извор]Наводи
[уреди | уреди извор]- ↑ „31. Ancient Life: Apex Chert Microfossils“. www.lpi.usra.edu. Посетено на 2022-03-12.
- ↑ Fredrickson, James K.; Zachara, John M.; Balkwill, David L.; Kennedy, David; Li, Shu-mei W.; Kostandarithes, Heather M.; Daly, Michael J.; Romine, Margaret F.; Brockman, Fred J. (2004-07-01). „Geomicrobiology of High-Level Nuclear Waste-Contaminated Vadose Sediments at the Hanford Site, Washington State“. Applied and Environmental Microbiology (англиски). 70 (7): 4230–4241. doi:10.1128/AEM.70.7.4230-4241.2004. ISSN 0099-2240. PMID 15240306. Архивирано од изворникот на 2018-01-03. Посетено на 2018-01-14.
- ↑ Rappé, Michael S.; Giovannoni, Stephen J. (2003-10-01). „The Uncultured Microbial Majority“. Annual Review of Microbiology. 57 (1): 369–394. doi:10.1146/annurev.micro.57.030502.090759. ISSN 0066-4227. Архивирано од изворникот на 2020-03-02. Посетено на 2018-01-14.
- ↑ „Những vi khuẩn gây bệnh thường gặp trong y học“. Đại học Dược Hà Nội (виетнамски). 2017-04-29. Посетено на 2018-06-28.
- ↑ Whitman, William B.; Coleman, David C.; Wiebe, William J. (1998-06-09). „Prokaryotes: The unseen majority“. Proceedings of the National Academy of Sciences (англиски). 95 (12): 6578–6583. ISSN 0027-8424. PMID 9618454. Архивирано од изворникот на 2011-10-30. Посетено на 2018-01-14.
- ↑ „Bacteria“. 2011-05-11. Архивирано од изворникот на 2011-05-11. Посетено на 2018-01-13.CS1-одржување: бот: непознат статус на изворната URL (link)
- ↑ Forbes, Shari. Decomposition Chemistry in a Burial Environment (англиски). стр. 203–223. doi:10.1201/9781420069921.ch8.[мртва врска]
- ↑ „Microbes Thrive in Deepest Spot on Earth“. Live Science. Посетено на 2018-01-13.
- ↑ Glud, Ronnie N.; Wenzhöfer, Frank; Middelboe, Mathias; Oguri, Kazumasa; Turnewitsch, Robert; Canfield, Donald E.; Kitazato, Hiroshi (април 2013). „High rates of microbial carbon turnover in sediments in the deepest oceanic trench on Earth“. Nature Geoscience (англиски). 6 (4): 284–288. doi:10.1038/ngeo1773. ISSN 1752-0908.
- ↑ „Intraterrestrials: Life Thrives in Ocean Floor“. Live Science. Посетено на 2018-01-13.
- ↑ Sender, Ron; Fuchs, Shai; Milo, Ron (2016-01-06). „Revised estimates for the number of human and bacteria cells in the body“. bioRxiv (англиски): 036103. doi:10.1101/036103.
- ↑ „A dynamic partnership: Celebrating our gut flora“. Anaerobe. 11 (5): 247–251. 2005-10-01. doi:10.1016/j.anaerobe.2005.05.001. ISSN 1075-9964.
- ↑ „WHO |“. www.who.int. Архивирано од изворникот на 2017-11-05. Посетено на 2018-01-13.
- ↑ „Metal-mining bacteria are green chemists“. ScienceDaily (англиски). Посетено на 2018-01-13.
- ↑ „Whole organism biocatalysis“. Current Opinion in Chemical Biology. 9 (2): 174–180. 2005-04-01. doi:10.1016/j.cbpa.2005.02.001. ISSN 1367-5931.
- ↑ 16,0 16,1 16,2 Woese, C. R.; Kandler, O.; Wheelis, M. L. (1990-06-01). „Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya“. Proceedings of the National Academy of Sciences (англиски). 87 (12): 4576–4579. doi:10.1073/pnas.87.12.4576. ISSN 0027-8424. PMID 2112744.
- ↑ „Henry George Liddell, Robert Scott, A Greek-English Lexicon, βακτήρ-ιον“. www.perseus.tufts.edu. Посетено на 2018-01-13.
- ↑ „Henry George Liddell, Robert Scott, A Greek-English Lexicon, βακτηρ-ία“. www.perseus.tufts.edu. Посетено на 2018-01-13.
- ↑ „bacteria | Origin and meaning of bacteria by Online Etymology Dictionary“. www.etymonline.com (англиски). Посетено на 2018-01-13.
- ↑ Schopf, J. W. (1994-07-19). „Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic“. Proceedings of the National Academy of Sciences (англиски). 91 (15): 6735–6742. ISSN 0027-8424. PMID 8041691. Архивирано од изворникот на 2014-08-23. Посетено на 2018-01-14.
- ↑ DeLong, Edward F.; Pace, Norman R. (2001-08-01). „Environmental Diversity of Bacteria and Archaea“. Systematic Biology. 50 (4): 470–478. doi:10.1080/10635150118513.
- ↑ Caetano-Anollés, Gustavo; Caetano-Anollés, Derek (July 2003). „An evolutionarily structured universe of protein architecture“. Genome Research. 13 (7): 1563–1571. doi:10.1101/gr.1161903. ISSN 1088-9051. PMID 12840035.
- ↑ Wang, Minglei; Yafremava, Liudmila S.; Caetano-Anollés, Derek; Mittenthal, Jay E.; Caetano-Anollés, Gustavo (November 2007). „Reductive evolution of architectural repertoires in proteomes and the birth of the tripartite world“. Genome Research. 17 (11): 1572–1585. doi:10.1101/gr.6454307. ISSN 1088-9051. PMC 2045140. PMID 17908824.CS1-одржување: PMC-формат (link)
- ↑ Giulio, Massimo Di (2003-12-01). „The Universal Ancestor and the Ancestor of Bacteria Were Hyperthermophiles“. Journal of Molecular Evolution (англиски). 57 (6): 721–730. doi:10.1007/s00239-003-2522-6. ISSN 0022-2844.
- ↑ Battistuzzi, Fabia U.; Feijao, Andreia; Hedges, S. Blair (2004-11-09). „A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land“. BMC Evolutionary Biology. 4: 44. doi:10.1186/1471-2148-4-44. ISSN 1471-2148.
- ↑ Poole, Anthony M.; Penny, David (2007-01-01). „Evaluating hypotheses for the origin of eukaryotes“. BioEssays (англиски). 29 (1): 74–84. doi:10.1002/bies.20516. ISSN 1521-1878.
- ↑ Dyall, Sabrina D.; Brown, Mark T.; Johnson, Patricia J. (2004-04-09). „Ancient Invasions: From Endosymbionts to Organelles“. Science (англиски). 304 (5668): 253–257. doi:10.1126/science.1094884. ISSN 0036-8075. PMID 15073369.
- ↑ „Endosymbiosis and evolution of the plant cell“. Current Opinion in Plant Biology. 2 (6): 513–519. 1999-12-01. doi:10.1016/S1369-5266(99)00025-4. ISSN 1369-5266.
- ↑ Schulz, Heide N.; Jørgensen, Bo Barker (2001-10-01). „Big Bacteria“. Annual Review of Microbiology. 55 (1): 105–137. doi:10.1146/annurev.micro.55.1.105. ISSN 0066-4227. Архивирано од изворникот на 2020-02-22. Посетено на 2018-01-14.
- ↑ 30,0 30,1 „Who are you calling simple?“. New Scientist. 211 (2821): 38–41. 2011-07-16. doi:10.1016/S0262-4079(11)61709-0. ISSN 0262-4079.
- ↑ Robertson, J; Gomersall, M; Gill, P (November 1975). „Mycoplasma hominis: growth, reproduction, and isolation of small viable cells“. Journal of Bacteriology. 124 (2): 1007–1018. ISSN 0021-9193. PMID 1102522.
- ↑ Velimirov, Branko (2001). „Nanobacteria, Ultramicrobacteria and Starvation Forms: A Search for the Smallest Metabolizing Bacterium“. Microbes and environments (англиски). 16 (2): 67–77. doi:10.1264/jsme2.2001.67. ISSN 1342-6311.
- ↑ Yang, Desirée C.; Blair, Kris M.; Salama, Nina R. (2016-03-01). „Staying in Shape: the Impact of Cell Shape on Bacterial Survival in Diverse Environments“. Microbiology and Molecular Biology Reviews (англиски). 80 (1): 187–203. doi:10.1128/MMBR.00031-15. ISSN 1092-2172. PMID 26864431.
- ↑ Cabeen, Matthew T.; Jacobs-Wagner, Christine (август 2005). „Bacterial cell shape“. Nature Reviews Microbiology (англиски). 3 (8): 601–610. doi:10.1038/nrmicro1205. ISSN 1740-1534.
- ↑ Young, Kevin D. (2006-09-01). „The Selective Value of Bacterial Shape“. Microbiology and Molecular Biology Reviews (англиски). 70 (3): 660–703. doi:10.1128/MMBR.00001-06. ISSN 1092-2172. PMID 16959965.
- ↑ Claessen, Dennis; Rozen, Daniel E.; Kuipers, Oscar P.; Søgaard-Andersen, Lotte; Wezel, Gilles P. van (февруари 2014). „Bacterial solutions to multicellularity: a tale of biofilms, filaments and fruiting bodies“. Nature Reviews Microbiology (англиски). 12 (2): 115–124. doi:10.1038/nrmicro3178. ISSN 1740-1534.
- ↑ Shimkets, Lawrence J. (1999-10-01). „Intercellular Signaling During Fruiting-Body Development of Myxococcus xanthus“. Annual Review of Microbiology. 53 (1): 525–549. doi:10.1146/annurev.micro.53.1.525. ISSN 0066-4227. Архивирано од изворникот на 2020-03-17. Посетено на 2018-01-14.
- ↑ Kaiser, Dale (2004-09-13). „Signaling in myxobacteria“. Annual Review of Microbiology. 58 (1): 75–98. doi:10.1146/annurev.micro.58.030603.123620. ISSN 0066-4227. Архивирано од изворникот на 2020-03-10. Посетено на 2018-01-14.
- ↑ 39,0 39,1 Donlan, Rodney M. „Biofilms: Microbial Life on Surfaces“. Emerging Infectious Diseases (англиски). 8 (9): 881–890. doi:10.3201/eid0809.020063. Архивирано од изворникот на 2017-12-09. Посетено на 2018-01-14.
- ↑ 40,0 40,1 Davey, Mary Ellen; O'toole, George A. (2000-12-01). „Microbial Biofilms: from Ecology to Molecular Genetics“. Microbiology and Molecular Biology Reviews (англиски). 64 (4): 847–867. doi:10.1128/MMBR.64.4.847-867.2000. ISSN 1092-2172. PMID 11104821.
- ↑ Donlan, Rodney M.; Costerton, J. William (2002-04-01). „Biofilms: Survival Mechanisms of Clinically Relevant Microorganisms“. Clinical Microbiology Reviews (англиски). 15 (2): 167–193. doi:10.1128/CMR.15.2.167-193.2002. ISSN 0893-8512. PMID 11932229. Архивирано од изворникот на 2018-06-19. Посетено на 2018-01-14.
- ↑ 42,0 42,1 L., Slonczewski, Joan (2013). Microbiology : an evolving science. [S.l.]: W W Norton. ISBN 9780393123685. OCLC 956340090.
- ↑ Bobik, Thomas A. (2006-05-01). „Polyhedral organelles compartmenting bacterial metabolic processes“. Applied Microbiology and Biotechnology (англиски). 70 (5): 517–525. doi:10.1007/s00253-005-0295-0. ISSN 0175-7598.
- ↑ Yeates, Todd O.; Kerfeld, Cheryl A.; Heinhorst, Sabine; Cannon, Gordon C.; Shively, Jessup M. (септември 2008). „Protein-based organelles in bacteria: carboxysomes and related microcompartments“. Nature Reviews Microbiology (англиски). 6 (9): 681–691. doi:10.1038/nrmicro1913. ISSN 1740-1534.
- ↑ Kerfeld, Cheryl A.; Sawaya, Michael R.; Tanaka, Shiho; Nguyen, Chau V.; Phillips, Martin; Beeby, Morgan; Yeates, Todd O. (2005-08-05). „Protein Structures Forming the Shell of Primitive Bacterial Organelles“. Science (англиски). 309 (5736): 936–938. doi:10.1126/science.1113397. ISSN 0036-8075. PMID 16081736.
- ↑ Gitai, Zemer (2005-03-11). „The New Bacterial Cell Biology: Moving Parts and Subcellular Architecture“. Cell (англиски). 120 (5): 577–586. doi:10.1016/j.cell.2005.02.026. ISSN 0092-8674.
- ↑ Shih, Yu-Ling; Rothfield, Lawrence (2006-09-01). „The Bacterial Cytoskeleton“. Microbiology and Molecular Biology Reviews (англиски). 70 (3): 729–754. doi:10.1128/MMBR.00017-06. ISSN 1092-2172. PMID 16959967.
- ↑ Norris, Vic; Blaauwen, Tanneke den; Cabin-Flaman, Armelle; Doi, Roy H.; Harshey, Rasika; Janniere, Laurent; Jimenez-Sanchez, Alfonso; Jin, Ding Jun; Levin, Petra Anne (2007-03-01). „Functional Taxonomy of Bacterial Hyperstructures“. Microbiology and Molecular Biology Reviews (англиски). 71 (1): 230–253. doi:10.1128/MMBR.00035-06. ISSN 1092-2172. PMID 17347523.
- ↑ Harold, F M (June 1972). „Conservation and transformation of energy by bacterial membranes“. Bacteriological Reviews. 36 (2): 172–230. ISSN 0005-3678. PMID 4261111.
- ↑ Bryant, Donald A.; Frigaard, Niels-Ulrik (2006-11-01). „Prokaryotic photosynthesis and phototrophy illuminated“. Trends in Microbiology (англиски). 14 (11): 488–496. doi:10.1016/j.tim.2006.09.001. ISSN 0966-842X. PMID 16997562.
- ↑ Pšenčík, J.; Ikonen, T.P.; Laurinmäki, P.; Merckel, M.C.; Butcher, S.J.; Serimaa, R.E.; Tuma, R. (2004-08-01). „Lamellar Organization of Pigments in Chlorosomes, the Light Harvesting Complexes of Green Photosynthetic Bacteria“. Biophysical Journal (англиски). 87 (2): 1165–1172. doi:10.1529/biophysj.104.040956. ISSN 0006-3495.
- ↑ Thanbichler, Martin; Wang, Sherry C.; Shapiro, Lucy (2005-10-15). „The bacterial nucleoid: A highly organized and dynamic structure“. Journal of Cellular Biochemistry (англиски). 96 (3): 506–521. doi:10.1002/jcb.20519. ISSN 1097-4644.
- ↑ Poehlsgaard, Jacob; Douthwaite, Stephen (ноември 2005). „The bacterial ribosome as a target for antibiotics“. Nature Reviews Microbiology (англиски). 3 (11): 870–881. doi:10.1038/nrmicro1265. ISSN 1740-1534.
- ↑ Yeo, Marcus; Chater, Keith (2005). „The interplay of glycogen metabolism and differentiation provides an insight into the developmental biology of Streptomyces coelicolor“. Microbiology. 151 (3): 855–861. doi:10.1099/mic.0.27428-0.
- ↑ Shiba, T.; Tsutsumi, K.; Ishige, K.; Noguchi, T. (March 2000). „Inorganic polyphosphate and polyphosphate kinase: their novel biological functions and applications“. Biochemistry. Biokhimiia. 65 (3): 315–323. ISSN 0006-2979. PMID 10739474.
- ↑ Brune, Daniel C. (1995-06-01). „Isolation and characterization of sulfur globule proteins from Chromatium vinosum and Thiocapsa roseopersicina“. Archives of Microbiology (англиски). 163 (6): 391–399. doi:10.1007/BF00272127. ISSN 0302-8933.
- ↑ Kadouri, Daniel; Jurkevitch, Edouard; Okon, Yaacov; Castro-Sowinski, Susana (2005-01-01). „Ecological and Agricultural Significance of Bacterial Polyhydroxyalkanoates“. Critical Reviews in Microbiology. 31 (2): 55–67. doi:10.1080/10408410590899228. ISSN 1040-841X. PMID 15986831.
- ↑ Walsby, A E (March 1994). „Gas vesicles“. Microbiological Reviews. 58 (1): 94–144. ISSN 0146-0749. PMID 8177173.
- ↑ van, Heijenoort, Jean (2001-03-01). „Formation of the glycan chains in the synthesis of bacterial peptidoglycan“. Glycobiology (англиски). 11 (3). doi:10.1093/glycob/11.3.25R. ISSN 0959-6658.
- ↑ 60,0 60,1 Koch, Arthur L. (2003-10-01). „Bacterial Wall as Target for Attack Past, Present, and Future Research“. Clinical Microbiology Reviews (англиски). 16 (4): 673–687. doi:10.1128/CMR.16.4.673-687.2003. ISSN 0893-8512. PMID 14557293. Архивирано од изворникот на 2018-06-01. Посетено на 2018-01-14.
- ↑ Gram, HC (1884). „\"{U}ber die isolierte F\"{a}rbung der Schizomyceten in Schnitt- und Trockenpr\"{a}paraten“. Fortschritte der Medizin. 2: 185–89.
- ↑ Hugenholtz, Philip (2002-01-29). „Exploring prokaryotic diversity in the genomic era“. Genome Biology. 3: reviews0003. doi:10.1186/gb-2002-3-2-reviews0003. ISSN 1474-760X.
- ↑ „Microbiology and drug resistance mechanisms of fully resistant pathogens“. Current Opinion in Microbiology. 7 (5): 439–444. 2004-10-01. doi:10.1016/j.mib.2004.08.007. ISSN 1369-5274.
- ↑ Alderwick, Luke J.; Harrison, James; Lloyd, Georgina S.; Birch, Helen L. (2015-08-01). „The Mycobacterial Cell Wall—Peptidoglycan and Arabinogalactan“. Cold Spring Harbor Perspectives in Medicine (англиски). 5 (8): a021113. doi:10.1101/cshperspect.a021113. ISSN 2157-1422. PMID 25818664.
- ↑ „Structural Research on Surface Layers: A Focus on Stability, Surface Layer Homology Domains, and Surface Layer–Cell Wall Interactions“. Journal of Structural Biology. 124 (2–3): 276–302. 1998-12-15. doi:10.1006/jsbi.1998.4070. ISSN 1047-8477.
- ↑ Beveridge, T. J.; Pouwels, P. H.; Sára, M.; Kotiranta, A.; Lounatmaa, K.; Kari, K.; Kerosuo, E.; Haapasalo, M.; Egelseer, E. M. (June 1997). „Functions of S-layers“. FEMS microbiology reviews. 20 (1–2): 99–149. ISSN 0168-6445. PMID 9276929.
- ↑ „The Bacterial Flagellar Motor: Structure and Function of a Complex Molecular Machine“. 233. 2004-01-01: 93–134. doi:10.1016/S0074-7696(04)33003-2. ISSN 0074-7696. Наводот journal бара
|journal=(help) - ↑ Beachey, E. H. (1981-03-01). „Bacterial Adherence: Adhesin-Receptor Interactions Mediating the Attachment of Bacteria to Mucosal Surfaces“. Journal of Infectious Diseases (англиски). 143 (3): 325–345. doi:10.1093/infdis/143.3.325. ISSN 0022-1899.
- ↑ Silverman, Philip M. (1997-01-01). „Towards a structural biology of bacterial conjugation“. Molecular Microbiology (англиски). 23 (3): 423–429. doi:10.1046/j.1365-2958.1997.2411604.x. ISSN 1365-2958.
- ↑ Costa, Tiago R. D.; Felisberto-Rodrigues, Catarina; Meir, Amit; Prevost, Marie S.; Redzej, Adam; Trokter, Martina; Waksman, Gabriel (јуни 2015). „Secretion systems in Gram-negative bacteria: structural and mechanistic insights“. Nature Reviews Microbiology (англиски). 13 (6): 343–359. doi:10.1038/nrmicro3456. ISSN 1740-1534.
- ↑ Stokes, Richard W.; Norris-Jones, Raymond; Brooks, Donald E.; Beveridge, Terry J.; Doxsee, Dan; Thorson, Lisa M. (2004-10-01). „The Glycan-Rich Outer Layer of the Cell Wall of Mycobacterium tuberculosis Acts as an Antiphagocytic Capsule Limiting the Association of the Bacterium with Macrophages“. Infection and Immunity (англиски). 72 (10): 5676–5686. doi:10.1128/IAI.72.10.5676-5686.2004. ISSN 0019-9567. PMID 15385466. Архивирано од изворникот на 2018-02-27. Посетено на 2018-01-14.
- ↑ „The capsule of Mycobacterium tuberculosis and its implications for pathogenicity“. Tubercle and Lung Disease. 79 (3): 153–169. 1999-06-01. doi:10.1054/tuld.1998.0200. ISSN 0962-8479.
- ↑ Finlay, B. B.; Falkow, S. (June 1997). „Common themes in microbial pathogenicity revisited“. Microbiology and molecular biology reviews: MMBR. 61 (2): 136–169. ISSN 1092-2172. PMID 9184008.
- ↑ Nicholson, Wayne L.; Munakata, Nobuo; Horneck, Gerda; Melosh, Henry J.; Setlow, Peter (2000-09-01). „Resistance of Bacillus Endospores to Extreme Terrestrial and Extraterrestrial Environments“. Microbiology and Molecular Biology Reviews (англиски). 64 (3): 548–572. doi:10.1128/MMBR.64.3.548-572.2000. ISSN 1092-2172. PMID 10974126.
- ↑ 75,0 75,1 McKenney, Peter T.; Driks, Adam; Eichenberger, Patrick (јануари 2013). „The Bacillus subtilis endospore: assembly and functions of the multilayered coat“. Nature Reviews Microbiology (англиски). 11 (1): 33–44. doi:10.1038/nrmicro2921. ISSN 1740-1534.
- ↑ Nicholson, Wayne L.; Fajardo-Cavazos, Patricia; Rebeil, Roberto; Slieman, Tony A.; Riesenman, Paul J.; Law, Jocelyn F.; Xue, Yaming (2002-12-01). „Bacterial endospores and their significance in stress resistance“. Antonie van Leeuwenhoek (англиски). 81 (1–4): 27–32. doi:10.1023/A:1020561122764. ISSN 0003-6072.
- ↑ Vreeland, Russell H.; Rosenzweig, William D.; Powers, Dennis W. (октомври 2000). „Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal“. Nature (англиски). 407 (6806): 897–900. doi:10.1038/35038060. ISSN 1476-4687.
- ↑ Cano, R. J.; Borucki, M. K. (1995-05-19). „Revival and identification of bacterial spores in 25- to 40-million-year-old Dominican amber“. Science (англиски). 268 (5213): 1060–1064. doi:10.1126/science.7538699. ISSN 0036-8075. PMID 7538699.
- ↑ „The solar UV environment and bacterial spore UV resistance: considerations for Earth-to-Mars transport by natural processes and human spaceflight“. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis. 571 (1–2): 249–264. 2005-04-01. doi:10.1016/j.mrfmmm.2004.10.012. ISSN 0027-5107.
- ↑ Hatheway, C L (January 1990). „Toxigenic clostridia“. Clinical Microbiology Reviews. 3 (1): 66–98. ISSN 0893-8512. PMID 2404569.
- ↑ Nealson, Kenneth H. (1999-01-01). „Post-Viking Microbiology: New Approaches, New Data, New Insights“. Origins of life and evolution of the biosphere (англиски). 29 (1): 73–93. doi:10.1023/A:1006515817767. ISSN 0169-6149.
- ↑ Xu, Jianping (2006-06-01). „INVITED REVIEW: Microbial ecology in the age of genomics and metagenomics: concepts, tools, and recent advances“. Molecular Ecology (англиски). 15 (7): 1713–1731. doi:10.1111/j.1365-294X.2006.02882.x. ISSN 1365-294X.
- ↑ „Comparative biochemistry of Archaea and Bacteria“. Current Opinion in Genetics & Development. 1 (4): 544–551. 1991-12-01. doi:10.1016/S0959-437X(05)80206-0. ISSN 0959-437X.
- ↑ Hellingwerf, K. J.; Crielaard, W.; Hoff, W. D.; Matthijs, H. C. P.; Mur, L. R.; Rotterdam, B. J. van (1994-12-01). „Photobiology of Bacteria“. Antonie van Leeuwenhoek (англиски). 65 (4): 331–347. doi:10.1007/BF00872217. ISSN 0003-6072.
- ↑ Seitzinger, S.; Harrison, J. A.; Böhlke, J. K.; Bouwman, A. F.; Lowrance, R.; Peterson, B.; Tobias, C.; Drecht, G. Van (2006-12-01). „Denitrification Across Landscapes and Waterscapes: A Synthesis“. Ecological Applications (англиски). 16 (6): 2064–2090. doi:10.1890/1051-0761(2006)016[2064:DALAWA]2.0.CO;2. ISSN 1939-5582.
- ↑ Zumft, W. G. (December 1997). „Cell biology and molecular basis of denitrification“. Microbiology and molecular biology reviews: MMBR. 61 (4): 533–616. ISSN 1092-2172. PMID 9409151.
- ↑ Acetogenesis | SpringerLink (англиски). doi:10.1007/978-1-4615-1777-1.
- ↑ Drake, Harold L.; Daniel, Steven L.; Küsel, Kirsten; Matthies, Carola; Kuhner, Carla; Braus-Stromeyer, Susanna (1997-01-01). „Acetogenic bacteria: what are the in situ consequences of their diverse metabolic versatilities?“. BioFactors (англиски). 6 (1): 13–24. doi:10.1002/biof.5520060103. ISSN 1872-8081.
- ↑ „Bacterial Fermentation“. www.els.net (англиски). Архивирано од изворникот на 2018-01-22. Посетено на 2018-01-14.
- ↑ Belser, L W (1979-10-01). „Population Ecology of Nitrifying Bacteria“. Annual Review of Microbiology. 33 (1): 309–333. doi:10.1146/annurev.mi.33.100179.001521. ISSN 0066-4227. Архивирано од изворникот на 2019-12-27. Посетено на 2018-01-14.
- ↑ Andrews, Simon; Norton, Ian; Salunkhe, Arvindkumar S.; Goodluck, Helen; Aly, Wafaa S. M.; Mourad-Agha, Hanna; Cornelis, Pierre (2013). Metallomics and the Cell. Metal Ions in Life Sciences (англиски). Springer, Dordrecht. стр. 203–239. doi:10.1007/978-94-007-5561-1_7. ISBN 9789400755604.
- ↑ Dalton, Howard (2005-06-29). „The Leeuwenhoek Lecture 2000 The natural and unnatural history of methane-oxidizing bacteria“. Philosophical Transactions of the Royal Society of London B: Biological Sciences (англиски). 360 (1458): 1207–1222. doi:10.1098/rstb.2005.1657. ISSN 0962-8436. PMID 16147517.
- ↑ Zehr, Jonathan P.; Jenkins, Bethany D.; Short, Steven M.; Steward, Grieg F. (2003-07-01). „Nitrogenase gene diversity and microbial community structure: a cross-system comparison“. Environmental Microbiology (англиски). 5 (7): 539–554. doi:10.1046/j.1462-2920.2003.00451.x. ISSN 1462-2920.
- ↑ Lonhienne, Thierry G. A.; Sagulenko, Evgeny; Webb, Richard I.; Lee, Kuo-Chang; Franke, Josef; Devos, Damien P.; Nouwens, Amanda; Carroll, Bernard J.; Fuerst, John A. (2010-07-20). „Endocytosis-like protein uptake in the bacterium Gemmata obscuriglobus“. Proceedings of the National Academy of Sciences (англиски). 107 (29): 12883–12888. doi:10.1073/pnas.1001085107. ISSN 0027-8424. PMID 20566852.
- ↑ Koch, Arthur L. (2002-01-01). „Control of the Bacterial Cell Cycle by Cytoplasmic Growth“. Critical Reviews in Microbiology. 28 (1): 61–77. doi:10.1080/1040-840291046696. ISSN 1040-841X.
- ↑ Eagon, R. G. (April 1962). „PSEUDOMONAS NATRIEGENS, A MARINE BACTERIUM WITH A GENERATION TIME OF LESS THAN 10 MINUTES“. Journal of Bacteriology. 83 (4): 736–737. ISSN 0021-9193. PMID 13888946.
- ↑ „LABORATORY DIAGNOSIS OF CENTRAL NERVOUS SYSTEM INFECTIONS“. Infectious Disease Clinics of North America. 15 (4): 1047–1071. 2001-12-01. doi:10.1016/S0891-5520(05)70186-0. ISSN 0891-5520.
- ↑ Challis, Gregory L.; Hopwood, David A. (2003-11-25). „Synergy and contingency as driving forces for the evolution of multiple secondary metabolite production by Streptomyces species“. Proceedings of the National Academy of Sciences (англиски). 100 (suppl 2): 14555–14561. doi:10.1073/pnas.1934677100. ISSN 0027-8424. PMID 12970466.
- ↑ Kooijman, S. a. L. M.; Auger, P.; Poggiale, J. C.; Kooi, B. W. (2003-08-01). „Quantitative steps in symbiogenesis and the evolution of homeostasis“. Biological Reviews (англиски). 78 (3): 435–463. doi:10.1017/S1464793102006127. ISSN 1469-185X.
- ↑ „Individual-based modelling of bacterial cultures to study the microscopic causes of the lag phase“. Journal of Theoretical Biology. 241 (4): 939–953. 2006-08-21. doi:10.1016/j.jtbi.2006.01.029. ISSN 0022-5193.
- ↑ „General stress response of Bacillus subtilis and other bacteria“. 44. 2001-01-01: 35–91. doi:10.1016/S0065-2911(01)44011-2. ISSN 0065-2911. Наводот journal бара
|journal=(help) - ↑ „JGI GOLD | Home“. gold.jgi.doe.gov. Посетено на 2018-01-14.
- ↑ Nakabachi, Atsushi; Yamashita, Atsushi; Toh, Hidehiro; Ishikawa, Hajime; Dunbar, Helen E.; Moran, Nancy A.; Hattori, Masahira (2006-10-13). „The 160-Kilobase Genome of the Bacterial Endosymbiont Carsonella“. Science (англиски). 314 (5797): 267–267. doi:10.1126/science.1134196. ISSN 0036-8075. PMID 17038615.
- ↑ Pradella, Silke; Hans, Astrid; Spröer, Cathrin; Reichenbach, Hans; Gerth, Klaus; Beyer, Stefan (2002-12-01). „Characterisation, genome size and genetic manipulation of the myxobacterium Sorangium cellulosum So ce56“. Archives of Microbiology (англиски). 178 (6): 484–492. doi:10.1007/s00203-002-0479-2. ISSN 0302-8933.
- ↑ Belfort, M; Reaban, M E; Coetzee, T; Dalgaard, J Z (July 1995). „Prokaryotic introns and inteins: a panoply of form and function“. Journal of Bacteriology. 177 (14): 3897–3903. ISSN 0021-9193. PMID 7608058.
- ↑ 106,0 106,1 „Management of multipartite genomes: the Vibrio cholerae model“. Current Opinion in Microbiology. 22: 120–126. 2014-12-01. doi:10.1016/j.mib.2014.10.003. ISSN 1369-5274.
- ↑ Hinnebusch, Joe; Tilly, Kit (1993-12-01). „Linear plasmids and chromosomes in bacteria“. Molecular Microbiology (англиски). 10 (5): 917–922. doi:10.1111/j.1365-2958.1993.tb00963.x. ISSN 1365-2958.
- ↑ Fraser, Claire M.; Casjens, Sherwood; Huang, Wai Mun; Sutton, Granger G.; Clayton, Rebecca; Lathigra, Raju; White, Owen; Ketchum, Karen A.; Dodson, Robert (декември 1997). „Genomic sequence of a Lyme disease spirochaete, Borrelia burgdorferi“. Nature (англиски). 390 (6660): 580–586. doi:10.1038/37551. ISSN 1476-4687.
- ↑ Lin, Y. S.; Kieser, H. M.; Hopwood, D. A.; Chen, C. W. (December 1993). „The chromosomal DNA of Streptomyces lividans 66 is linear“. Molecular Microbiology. 10 (5): 923–933. ISSN 0950-382X. PMID 7934869.
- ↑ Kado, Clarence I. (2014-10-10). „Historical Events That Spawned the Field of Plasmid Biology“. Microbiology Spectrum (англиски). 2 (5). doi:10.1128/microbiolspec.plas-0019-2013. ISSN 2165-0497.
- ↑ Baxter, Jamie C.; Funnell, Barbara E. (2014-11-07). „Plasmid Partition Mechanisms“. Microbiology Spectrum (англиски). 2 (6). doi:10.1128/microbiolspec.plas-0023-2014. ISSN 2165-0497.
- ↑ Denamur, Erick; Matic, Ivan (2006-05-01). „Evolution of mutation rates in bacteria“. Molecular Microbiology (англиски). 60 (4): 820–827. doi:10.1111/j.1365-2958.2006.05150.x. ISSN 1365-2958.
- ↑ Wright, Barbara E. (2004-05-01). „Stress-directed adaptive mutations and evolution“. Molecular Microbiology (англиски). 52 (3): 643–650. doi:10.1111/j.1365-2958.2004.04012.x. ISSN 1365-2958.
- ↑ Chen, Inês; Dubnau, David (март 2004). „DNA uptake during bacterial transformation“. Nature Reviews Microbiology (англиски). 2 (3): 241–249. doi:10.1038/nrmicro844. ISSN 1740-1534.
- ↑ „Who's competent and when: regulation of natural genetic competence in bacteria“. Trends in Genetics. 12 (4): 150–155. 1996-04-01. doi:10.1016/0168-9525(96)10014-7. ISSN 0168-9525.
- ↑ AKAMATSU, Takashi; TAGUCHI, Hisataka (2001-01-01). „Incorporation of the Whole Chromosomal DNA in Protoplast Lysates into Competent Cells of Bacillus subtilis“. Bioscience, Biotechnology, and Biochemistry. 65 (4): 823–829. doi:10.1271/bbb.65.823. ISSN 0916-8451. PMID 11388459.
- ↑ „Natural genetic transformation: prevalence, mechanisms and function“. Research in Microbiology. 158 (10): 767–778. 2007-12-01. doi:10.1016/j.resmic.2007.09.004. ISSN 0923-2508.
- ↑ „Adaptive value of sex in microbial pathogens“. Infection, Genetics and Evolution. 8 (3): 267–285. 2008-05-01. doi:10.1016/j.meegid.2008.01.002. ISSN 1567-1348.
- ↑ „Genetic Exchange between Bacteria in the Environment“. Plasmid. 42 (2): 73–91. 1999-09-01. doi:10.1006/plas.1999.1421. ISSN 0147-619X.
- ↑ Hastings, P. J.; Rosenberg, Susan M.; Slack, Andrew (2004-09-01). „Antibiotic-induced lateral transfer of antibiotic resistance“. Trends in Microbiology (англиски). 12 (9): 401–404. doi:10.1016/j.tim.2004.07.003. ISSN 0966-842X. PMID 15337159.
- ↑ Brüssow, Harald; Canchaya, Carlos; Hardt, Wolf-Dietrich (2004-09-01). „Phages and the Evolution of Bacterial Pathogens: from Genomic Rearrangements to Lysogenic Conversion“. Microbiology and Molecular Biology Reviews (англиски). 68 (3): 560–602. doi:10.1128/MMBR.68.3.560-602.2004. ISSN 1092-2172. PMID 15353570.
- ↑ Bickle, T. A.; Krüger, D. H. (June 1993). „Biology of DNA restriction“. Microbiological Reviews. 57 (2): 434–450. ISSN 0146-0749. PMID 8336674.
- ↑ Barrangou, Rodolphe; Fremaux, Christophe; Deveau, Hélène; Richards, Melissa; Boyaval, Patrick; Moineau, Sylvain; Romero, Dennis A.; Horvath, Philippe (2007-03-23). „CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes“. Science (англиски). 315 (5819): 1709–1712. doi:10.1126/science.1138140. ISSN 0036-8075. PMID 17379808.
- ↑ Brouns, Stan J. J.; Jore, Matthijs M.; Lundgren, Magnus; Westra, Edze R.; Slijkhuis, Rik J. H.; Snijders, Ambrosius P. L.; Dickman, Mark J.; Makarova, Kira S.; Koonin, Eugene V. (2008-08-15). „Small CRISPR RNAs Guide Antiviral Defense in Prokaryotes“. Science (англиски). 321 (5891): 960–964. doi:10.1126/science.1159689. ISSN 0036-8075. PMID 18703739.
- ↑ 125,0 125,1 125,2 Bardy, Sonia L.; Ng, Sandy Y. M.; Jarrell, Ken F. (2003). „Prokaryotic motility structures“. Microbiology. 149 (2): 295–304. doi:10.1099/mic.0.25948-0.
- ↑ Sheetz, Michael P.; Merz, Alexey J.; So, Magdalene (септември 2000). „Pilus retraction powers bacterial twitching motility“. Nature (англиски). 407 (6800): 98–102. doi:10.1038/35024105. ISSN 1476-4687.
- ↑ Macnab, Robert M. (December 1999). „The Bacterial Flagellum: Reversible Rotary Propellor and Type III Export Apparatus“. Journal of Bacteriology. 181 (23): 7149–7153. ISSN 0021-9193. PMID 10572114.
- ↑ Wu, Mingming; Roberts, John W.; Kim, Sue; Koch, Donald L.; DeLisa, Matthew P. (2006-07-01). „Collective Bacterial Dynamics Revealed Using a Three-Dimensional Population-Scale Defocused Particle Tracking Technique“. Applied and Environmental Microbiology (англиски). 72 (7): 4987–4994. doi:10.1128/AEM.00158-06. ISSN 0099-2240. PMID 16820497. Архивирано од изворникот на 2018-06-01. Посетено на 2018-01-14.
- ↑ Lux, Renate; Shi, Wenyuan (2016-12-01). „Chemotaxis-guided Movements in Bacteria“. Critical Reviews in Oral Biology & Medicine (англиски). 15 (4): 207–220. doi:10.1177/154411130401500404.
- ↑ Goldberg, Marcia B. (2001-12-01). „Actin-Based Motility of Intracellular Microbial Pathogens“. Microbiology and Molecular Biology Reviews (англиски). 65 (4): 595–626. doi:10.1128/MMBR.65.4.595-626.2001. ISSN 1092-2172. PMID 11729265.
- ↑ „Wayback Machine“ (PDF). 2011-07-17. Архивирано од изворникот (PDF) на 2011-07-17. Посетено на 2018-01-14.
- ↑ Costerton, J W; Lewandowski, Z; Caldwell, D E; Korber, D R; Lappin-Scott, H M (1995-10-01). „Microbial Biofilms“. Annual Review of Microbiology. 49 (1): 711–745. doi:10.1146/annurev.mi.49.100195.003431. ISSN 0066-4227. Архивирано од изворникот на 2019-04-24. Посетено на 2018-01-14.
- ↑ Miller, Melissa B.; Bassler, Bonnie L. (2001-10-01). „Quorum Sensing in Bacteria“. Annual Review of Microbiology. 55 (1): 165–199. doi:10.1146/annurev.micro.55.1.165. ISSN 0066-4227. Архивирано од изворникот на 2019-04-05. Посетено на 2018-01-14.
- ↑ Boucher, Yan; Douady, Christophe J.; Papke, R. Thane; Walsh, David A.; Boudreau, Mary Ellen R.; Nesbø, Camilla L.; Case, Rebecca J.; Doolittle, W. Ford (2003-11-28). „Lateral Gene Transfer and the Origins of Prokaryotic Groups“. Annual Review of Genetics. 37 (1): 283–328. doi:10.1146/annurev.genet.37.050503.084247. ISSN 0066-4197. Архивирано од изворникот на 2019-02-16. Посетено на 2018-01-14.
- ↑ Olsen, G. J.; Woese, C. R.; Overbeek, R. A. (1996-03-01). „The Winds of (evolutionary) Change: Breathing New Life into Microbiology“ (англиски). Наводот journal бара
|journal=(help) - ↑ Gupta, Radhey S. (2000-01-01). „The Natural Evolutionary Relationships among Prokaryotes“. Critical Reviews in Microbiology. 26 (2): 111–131. doi:10.1080/10408410091154219. ISSN 1040-841X.
- ↑ „Evolutionary aspects of whole-genome biology“. Current Opinion in Structural Biology. 15 (3): 248–253. 2005-06-01. doi:10.1016/j.sbi.2005.04.001. ISSN 0959-440X.
- ↑ Woods, G L; Walker, D H (July 1996). „Detection of infection or infectious agents by use of cytologic and histologic stains“. Clinical Microbiology Reviews. 9 (3): 382–404. ISSN 0893-8512. PMID 8809467.
- ↑ Weinstein, M. P. (March 1994). „Clinical importance of blood cultures“. Clinics in Laboratory Medicine. 14 (1): 9–16. ISSN 0272-2712. PMID 8181237.
- ↑ Louie, Marie; Louie, Lisa; Simor, Andrew E. (2000-08-08). „The role of DNA amplification technology in the diagnosis of infectious diseases“. Canadian Medical Association Journal (англиски). 163 (3): 301–309. ISSN 0820-3946. PMID 10951731.
- ↑ Oliver, James D. (February 2005). „The viable but nonculturable state in bacteria“. Journal of Microbiology (Seoul, Korea). 43 Spec No: 93–100. ISSN 1225-8873. PMID 15765062.
- ↑ „Number of published names“. 2012-01-19. Архивирано од изворникот на 2012-01-19. Посетено на 2018-01-14.CS1-одржување: бот: непознат статус на изворната URL (link)
- ↑ Curtis, Thomas P.; Sloan, William T.; Scannell, Jack W. (2002-08-06). „Estimating prokaryotic diversity and its limits“. Proceedings of the National Academy of Sciences (англиски). 99 (16): 10494–10499. doi:10.1073/pnas.142680199. ISSN 0027-8424. PMID 12097644.
- ↑ Schloss, Patrick D.; Handelsman, Jo (2004-12-01). „Status of the Microbial Census“. Microbiology and Molecular Biology Reviews (англиски). 68 (4): 686–691. doi:10.1128/MMBR.68.4.686-691.2004. ISSN 1092-2172. PMID 15590780.
- ↑ Martin, Mark O. (September 2002). „Predatory prokaryotes: an emerging research opportunity“. Journal of Molecular Microbiology and Biotechnology. 4 (5): 467–477. ISSN 1464-1801. PMID 12432957.
- ↑ Velicer, Gregory J.; Stredwick, Kristina L. (2002-12-01). „Experimental social evolution with Myxococcus xanthus“. Antonie van Leeuwenhoek (англиски). 81 (1–4): 155. doi:10.1023/A:1020546130033. ISSN 0003-6072.
- ↑ Guerrero, Ricardo; Pedrós-Alió, Carlos; Esteve, Isabel; Mas, Jordi; Chase, David; Margulis, Lynn (1986-04-01). „Predatory prokaryotes: Predation and primary consumption evolved in bacteria“. Proceedings of the National Academy of Sciences (англиски). 83 (7): 2138–2142. ISSN 0027-8424. PMID 11542073.
- ↑ Stams, Alfons J. M.; De Bok, Frank A. M.; Plugge, Caroline M.; Van Eekert, Miriam H. A.; Dolfing, Jan; Schraa, Gosse (2006-03-01). „Exocellular electron transfer in anaerobic microbial communities“. Environmental Microbiology (англиски). 8 (3): 371–382. doi:10.1111/j.1462-2920.2006.00989.x. ISSN 1462-2920.
- ↑ Barea, José-Miguel; Pozo, María José; Azcón, Rosario; Azcón-Aguilar, Concepción (2005-07-01). „Microbial co-operation in the rhizosphere“. Journal of Experimental Botany (англиски). 56 (417): 1761–1778. doi:10.1093/jxb/eri197. ISSN 0022-0957.
- ↑ O'Hara, Ann M.; Shanahan, Fergus (2006-07-01). „The gut flora as a forgotten organ“. EMBO reports (англиски). 7 (7): 688–693. doi:10.1038/sj.embor.7400731. ISSN 1469-221X. PMID 16819463. Архивирано од изворникот на 2018-01-26. Посетено на 2018-01-14.
- ↑ Zoetendal, Erwin G.; Vaughan, Elaine E.; De Vos, Willem M. (2006-03-01). „A microbial world within us“. Molecular Microbiology (англиски). 59 (6): 1639–1650. doi:10.1111/j.1365-2958.2006.05056.x. ISSN 1365-2958.
- ↑ Salminen, Seppo J.; Gueimonde, Miguel; Isolauri, Erika (May 2005). „Probiotics that modify disease risk“. The Journal of Nutrition. 135 (5): 1294–1298. ISSN 0022-3166. PMID 15867327.
- ↑ Belland, Robert J.; Ouellette, Scot P.; Gieffers, Jens; Byrne, Gerald I. (2004-02-01). „Chlamydia pneumoniae and atherosclerosis“. Cellular Microbiology (англиски). 6 (2): 117–127. doi:10.1046/j.1462-5822.2003.00352.x. ISSN 1462-5822.
- ↑ Heise, E R (February 1982). „Diseases associated with immunosuppression“. Environmental Health Perspectives. 43: 9–19. ISSN 0091-6765. PMC 1568899. PMID 7037390.CS1-одржување: PMC-формат (link)
- ↑ Saiman, Lisa (2004-01-01). „Microbiology of early CF lung disease“. Paediatric Respiratory Reviews (англиски). 5. doi:10.1016/S1526-0542(04)90065-6. ISSN 1526-0542.
- ↑ Yonath, Ada; Bashan, Anat (2004-09-13). „Ribosomal Crystallography: Initiation, Peptide Bond Formation, and Amino Acid Polymerization are Hampered by Antibiotics“. Annual Review of Microbiology. 58 (1): 233–251. doi:10.1146/annurev.micro.58.030603.123822. ISSN 0066-4227. Архивирано од изворникот на 2020-03-12. Посетено на 2018-01-14.
- ↑ Khachatourians, G G (1998-11-03). „Agricultural use of antibiotics and the evolution and transfer of antibiotic-resistant bacteria“. CMAJ: Canadian Medical Association Journal. 159 (9): 1129–1136. ISSN 0820-3946. PMC 1229782. PMID 9835883.CS1-одржување: PMC-формат (link)
- ↑ Hagedorn, S; Kaphammer, B (1994-10-01). „Microbial Biocatalysis in the Generation of Flavor and Fragrance Chemicals“. Annual Review of Microbiology. 48 (1): 773–800. doi:10.1146/annurev.mi.48.100194.004013. ISSN 0066-4227. Архивирано од изворникот на 2020-03-21. Посетено на 2018-01-15.
- ↑ Cohen, Yehuda (2002-12-01). „Bioremediation of oil by marine microbial mats“. International Microbiology (англиски). 5 (4): 189–193. doi:10.1007/s10123-002-0089-5. ISSN 1139-6709.
- ↑ Neves, Luiz Carlos Martins Das; Miyamura, Tábata Taemi Miazaki Ohara; Moraes, Dante Augusto; Penna, Thereza Christina Vessoni; Converti, Attilio (2006-03-01). „Biofiltration methods for the removal of phenolic residues“. Applied Biochemistry and Biotechnology (англиски). 129 (1–3): 130–152. doi:10.1385/ABAB:129:1:130. ISSN 0273-2289.
- ↑ „Production of fine chemicals using biocatalysis“. Current Opinion in Biotechnology. 10 (6): 595–603. 1999-12-01. doi:10.1016/S0958-1669(99)00040-3. ISSN 0958-1669.
- ↑ Aronson, Arthur I.; Shai, Yechiel (2001-02-01). „WhyBacillus thuringiensisinsecticidal toxins are so effective: unique features of their mode of action“. FEMS Microbiology Letters (англиски). 195 (1): 1–8. doi:10.1111/j.1574-6968.2001.tb10489.x. ISSN 0378-1097.
- ↑ Bozsik, András (2006-07-01). „Susceptibility of adult Coccinella septempunctata (Coleoptera: Coccinellidae) to insecticides with different modes of action“. Pest Management Science (англиски). 62 (7): 651–654. doi:10.1002/ps.1221. ISSN 1526-4998.
- ↑ Serres, Margrethe H.; Gopal, Shuba; Nahum, Laila A.; Liang, Ping; Gaasterland, Terry; Riley, Monica (2001-08-20). „A functional update of the Escherichia coliK-12 genome“. Genome Biology. 2: research0035. doi:10.1186/gb-2001-2-9-research0035. ISSN 1474-760X.
- ↑ Walsh, Gary (2005-04-01). „Therapeutic insulins and their large-scale manufacture“. Applied Microbiology and Biotechnology (англиски). 67 (2): 151–159. doi:10.1007/s00253-004-1809-x. ISSN 0175-7598.
- ↑ Graumann, Klaus; Premstaller, Andreas (2006-02-01). „Manufacturing of recombinant therapeutic proteins in microbial systems“. Biotechnology Journal (англиски). 1 (2): 164–186. doi:10.1002/biot.200500051. ISSN 1860-7314.
- ↑ Porter, J R (June 1976). „Antony van Leeuwenhoek: tercentenary of his discovery of bacteria“. Bacteriological Reviews. 40 (2): 260–269. ISSN 0005-3678. PMID 786250.
- ↑ Leewenhoeck, Anthony (1684-01-20). „An abstract of a letter from Mr. Anthony Leevvenhoeck at Delft, dated Sep. 17. 1683. Containing some microscopical observations, about animals in the scurf of the teeth, the substance call'd worms in the nose, the cuticula consisting of scales“. Philosophical Transactions (англиски). 14 (159): 568–574. doi:10.1098/rstl.1684.0030. ISSN 0261-0523.
- ↑ Leeuwenhoek, Antoni Van (1701-01-01). „IV. Part of a letter from Mr Antony Van Leeuwenhoek, concerning the worms in Sheeps livers, Gants and animalcula in the excrements of Frogs“. Philosophical Transactions (англиски). 22 (261): 509–518. doi:10.1098/rstl.1700.0013. ISSN 0261-0523.
- ↑ Leeuwenhoek, Antoni Van (1703-01-01). „IV. Part of a letter from Mr Antony van Leeuwenhoek, F. R. S. concerning green weeds growing in water, and some animalcula found about them“. Philosophical Transactions (англиски). 23 (283): 1304–1311. doi:10.1098/rstl.1702.0042. ISSN 0261-0523.
- ↑ Breed, Robert S.; Conn, H. J. (мај 1936). „The Status of the Generic Term Bacterium Ehrenberg 1828*“. Journal of Bacteriology. 31 (5): 517–518. ISSN 0021-9193. PMID 16559906.
- ↑ „Pasteur's Papers on the Germ Theory“. biotech.law.lsu.edu. Посетено на 2018-02-19.
- ↑ „HIV causes AIDS: Koch's postulates fulfilled“. Current Opinion in Immunology (англиски). 8 (5): 613–618. 1996-10-01. doi:10.1016/S0952-7915(96)80075-6. ISSN 0952-7915.
- ↑ „The Nobel Prize in Physiology or Medicine 1905“. www.nobelprize.org. Посетено на 2018-02-19.
- ↑ Austrian, Robert (September 1960). „THE GRAM STAIN AND THE ETIOLOGY OF LOBAR PNEUMONIA, AN HISTORICAL NOTE1“. Bacteriological Reviews. 24 (3): 261–265. ISSN 0005-3678. PMID 13685217.
- ↑ „Paul Ehrlich - Biographical“. www.nobelprize.org. Посетено на 2018-02-19.
- ↑ „Ilya Mechnikov - Biographical“. www.nobelprize.org. Посетено на 2018-02-19.
Литература
[уреди | уреди извор]- Holt JC, Bergey DH (1994). Bergey's Manual of Determinative Bacteriology (9th ed.). Baltimore: Williams & Wilkins. ISBN 0-683-00603-7.
- Atlas RM (1995). Principles of microbiology. St. Louis: Mosby. ISBN 0-8016-7790-4.
- Alcamo IE (2001). Fundamentals of microbiology. Boston: Jones and Bartlett. ISBN 0-7637-1067-9.
- Ogunseitan OA (2005). Microbial Diversity: Form and Function in Prokaryotes. Wiley-Blackwell. ISBN 978-1-4051-4448-3.
- Shively JM (2006). Complex Intracellular Structures in Prokaryotes (Microbiology Monographs). Berlin: Springer. ISBN 3-540-32524-7.
- White D, Drummond J, Fuqua C (2011) The Physiology and Biochemistry of Prokaryotes (4th ed.) Oxford University Press. ISBN 019539304X
- Madigan MT, Bender KS, Buckley DH, Sattley WM, Stahl DA (2017). Brock Biology of Microorganisms (15th ed.). London: Pearson. ISBN 0134261925.
Надворешни врски
[уреди | уреди извор]| „Бактерии“ на Ризницата ? |
- „Бактерии“ — Енциклопедија на живиот свет

- Бактерии кои делуваат на житните и другите растенија Архивирано на 23 февруари 2018 г. (на анг.)
- Ажурирана номенклатура на прокариоти (на анг.)
- Родови во доменот бактерии (на анг.)
- Најголемите бактерии Архивирано на 16 април 2017 г. (на анг.)
- Видеа Архивирано на 18 февруари 2018 г. на бактерии како пливаат и се превртуваат. (на анг.)
- Планета на бактерии Архивирано на 22 јуни 2019 г. од Стивен Џеј Гулд. (на анг.)
- Онлајн учебник по бактериологија Архивирано на 13 септември 2008 г. (на анг.)
- Анимиран водич за структура на бактериска клетка Архивирано на 13 септември 2008 г. (на анг.)
- Онлајн соработка за идентификација на бактерии (на анг.)
- PATRIC, биоинформатички ресурсен центар за патогени бактерии, спонзориран од NIAID (на анг.)
- Интерактивен симулатор на бактериска хемотакса - мрежно место кое користи едноставни алгоритми за да ја симулира бактериската хемотакса. (на анг.)
- Меѓуклеточна комуникација кај бактерии - онлајн предавање на Bonnie Bassler и ТЕД: Откривање на чудесниот комуникациски систем кај бактериите. (на анг.)
|
|
| Статијата „Бактерии“ е избрана статија. Ве повикуваме и Вас да напишете и предложите избрана статија (останати избрани статии). |